TOPIC: Лингвистико-Волновой геном. Следующий шаг. №1
Лингвистико-Волновой геном. Следующий шаг. №1
29 Окт 2010 14:50 #331
ППГ написал(а):
Хайдук, как с домашним заданием по кодировке 20 объектов тройками из четырех букв?
Наверное не заметили, что домашнее задание выполнено постом #325: из 4 букв можно составить 4^3 = 64 триплета, что более, чем достаточно для кодирования 20+ аминокислот
Лингвистико-Волновой геном. Следующий шаг. №1
29 Окт 2010 16:28 #332
Хайдук написал(а):
Петр, Вы сами выдвигаете удобные Вам условия и потом им любуетесь
. Детям уже ясно, что кодовую функцию несут триплеты, а не дуплеты. Из 4 букв можно составить 4^3 = 64 триплета, что более, чем достаточно для кодирования 20+ аминокислот. Вася фсё объяснил, избыточность и многозначность не проблемы, код немного сложнее, чем волногонщикам хотелось бы. Лучше направьте свои усилия к выяснению 96% мусора мусор ли это на самом деле или в нем прячутся некоторые регуляторные жемчужины. Бросьте высосанные из пальцев красиво выглядевшие (как голограммы
) натурфилософские мифы и займитесь реальной работой, а не мистериозным великорусским шовинистическим бредом
У-тю-тю, как разошелся... А рано. За домашнее задание - двойка. Дневник показать родителям. Пусть придут в школу ЛВГ №1 имени меня.
Принимаю круглосуточно.
Что вычислили число триплетов - 64, это хорошо, иначе кол поставил бы. Теперь напрягите остатки мозгов, вспомните условия задачки - кодируют в триплетах только первые две буквы (Гамов был частично прав), третьи не участвуют в кодировании. Кодирование идет по схеме два из трех (Lagerkvist, 1978). Как теперь решите задачку? Не решите - на второй год.
Лингвистико-Волновой геном. Следующий шаг. №1
29 Окт 2010 16:42 #333
ППГ написал(а):
третьи не участвуют в кодировании.
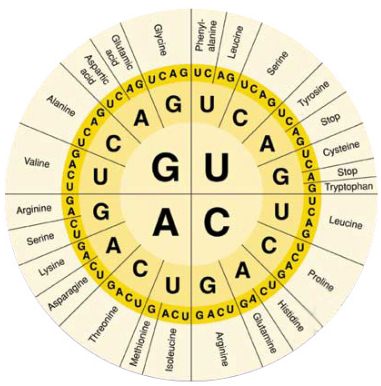
Впервые идея о существовании генетического кода сформулирована А. Дауном и Г. Гамовым в 1952-54, к-рые показали, что последовательность нуклеотидов, однозначно определяющая синтез той или иной аминокислоты, должна содержать не менее трех звеньев. Позднее было доказано, что такая последовательность состоит из трех нуклеотидов, названных кодоном, или триплетом. Т.к. молекулы нуклеиновых к-т, на к-рых происходит синтез мРНК или белка, состоят из остатков только четырех разных нуклеотидов, кодонов, отличающихся между собой, м. б. всего 64.
Все синтезируемые в процессе трансляции белки построены из остатков 20 аминокислот (т. наз. кодируемых). Какой именно кодон ответствен за включение той или иной аминокислоты, можно определить по таблице, в к-рой буквы А, Г, У, Ц обозначают основания, входящие в ну-клеотиды (соотв. аденин, гуанин, урацил, цитозин): в вертикальном ряду слева-в первый нуклеотид кодона, в горизонтальном ряду сверху-во второй, в вертикальном ряду справа-в третий. Трехбуквенные сочетания, напр. фен, сер, лей,-сокращенные назв. аминокислот. Прочерки в таблице означают, что три кодона-УАА, УАГ и УГА в нормальных условиях не кодируют к.-л. аминокислоты. Такие кодоны наз. бессмысленными, или нонсенс-кодонами. Они являются сигналами остановки синтеза полипептидной цепи.
В таблице представлены не все аминокислоты, встречающиеся в белках. В ней нет гидроксипролина и гидроксилизина, содержащихся в коллагене; фосфосерина-компонента всех фосфопротеидов; иодпроизводных тирозина, содержащихся в тиреоглобулине; цистина, к-рый часто встречается в белках, и нек-рых др. аминокислот. Все они-производные др. аминокислот, к-рые кодируются мРНК. Они образуются в результате модификации белков, происходящей после трансляции.
Генетический код специфичен: это означает, что каждый кодон кодирует только одну аминокислоту. Лишь два кодона, кодирующие валин (ГУГ) и метионин (АУГ), способны выполнять дополнит. ф-ции. Если они находятся в начале считываемой области мРНК, к ним присоединяется транспортная РНК (тРНК), несущая формилметионин, к-рый всегда находится в начале строящейся полипептидной цепи, а по завершении синтеза отщепляется целиком или отщепляет формильный остаток, превращаясь в остаток метионина. Т. обр., кодоны ГУГ и АУГ-инициаторы синтеза полипептидной цепи. Если же они не стоят первыми, то не отличаются по ф-циям от др. кодонов.
Генетический код называют вырожденным, поскольку 61 кодон кодирует всего 20 аминокислот. Поэтому почти каждой аминокислоте соответствует более чем один кодон. Вырожденность генетического кода неравномерна: для аргинина, серина и лейцина она шестикратна (т.е. для каждой из этих аминокислот имеется по шесть кодонов), тогда как для мн. др. аминокислот (тирозина, гистидина, фенилаланина и др.) лишь двукратна. Две аминокислоты (метионин и триптофан) представлены единств. кодонами. Кодоны-синонимы почти всегда отличаются друг от друга по последнему из трех нуклеотидов, тогда как первые два совпадают. Т. обр., код аминокислоты определяется в осн. первыми двумя буквами. Вырожденность генетического кода имеет важное значение для повышения устойчивости генетич. информации.
С механизмами трансляции связана еще одна особенность генетического кода: он неперекрывающийся. Кодоны транслируются всегда целиком; для кодирования невозможно использование элементов одного из них в сочетании с элементами соседнего. Рамкой, ограничивающей транслируемый кодон и перемещающейся скачком сразу на три нуклеотида, служит антикодон тРНК, к-рый представляет собой триплет нуклеотидов, комплементарный одному из кодонов и обусловливающий специфичность к нему. Т. обр., наблюдается линейное соответствие между последовательностью кодирующих триплетов и расположением остатков аминокислот в синтезируемом полипептиде, т.е. код имеет линейный непрерывающийся порядок считывания.
Важнейшее св-во генетического кода-его однонаправленность. Кодоны информативны только в том случае, если они считываются в одном направлении-от первого нуклеотида к последующим.
Генетический код универсален для всех живых существ. Возможны только небольшие видовые изменения, возникшие, вероятно, при эволюции и дифференцировке клеток. Большинство из них связано с вырожденностью кода и проявляется в преимуществ. использовании разных кодонов одной и той же аминокислоты и в различиях в структуре соответствующих тРНК в разных организмах или в разных тканях одного организма.
Представления об общих принципах и осн. св-вах генетического кода были разработаны Ф. Криком в 1957-65. Благодаря работам X. Кораны, X. Маттеи, М. Ниренберга и С. Очоа к 1966 был расшифрован нуклеотидный состав всех кодонов.
«ГЕНЕТИЧЕСКИЙ КОД»: И час М., Биологический код, пер. с англ., М., 1971. П. Л. Иванов.
Лингвистико-Волновой геном. Следующий шаг. №1
29 Окт 2010 17:53 #334
Забавно наблюдать зомбаж в действии. Люди, как под гипнозом. Никакие доводы не действуют. Логика молчит. Даже ссылки на сомнения самого автора модели белкового кода. Потратил почти 8 лет на молбиол.ру, чтобы хотя чуть задумались. Фантастика! Даже простейший пример не хотят видеть - триплет поли-уридин ДОЛЖЕН по канону кодировать ТОЛЬКО фенилаланин, а он, подлец такой, кодирует еще и лейцин. И вообще, кодоны ведут себя иногда просто неприлично - при аминокислотном голодании они запросто перекодируются, то же в присутствии некоторых антибиотиков, при теплошоке и пр. Да и кодов-то не один, а двадцать. Вот-те и универсальность... Словом, нехорошо как-то... Вот и пришли к ГМ-кушаньям... Кушайте на здоровье.
Лингвистико-Волновой геном. Следующий шаг. №1
29 Окт 2010 18:04 #335
ППГ написал(а):
Забавно наблюдать зомбаж в действии
Генетический код, система зашифровки наследственной информации в молекулах нуклеиновых кислот, реализующаяся у животных, растений, бактерий и вирусов в виде последовательности нуклеотидов. В природных нуклеиновых кислотах — дезоксирибонуклеиновой (ДНК) и рибонуклеиновой (РНК) — встречаются 5 распространённых типов нуклеотидов (по 4 в каждой нуклеиновой кислоте), различающихся по входящему в их состав азотистому основанию (см. Пуриновые основания, Пиримидиновые основания). В ДНК встречаются основания: аденин (А), гуанин (Г), цитозин (Ц), тимин (Т); в РНК вместо тимина присутствует урацил (У). Кроме них, в составе нуклеиновых кислот обнаружено около 20 редко встречающихся (т. н. неканонических, или минорных) оснований, а также необычных сахаров. Т. к. количество кодирующих знаков Г. к. (4) и число разновидностей аминокислот в белке (20) не совпадают, кодовое число (т. е. количество нуклеотидов, кодирующих 1 аминокислоту) не может быть равно 1. Различных сочетаний по 2 нуклеотида возможно лишь 42=16, но этого также недостаточно для зашифровки всех аминокислот. Американский учёный Г. Гамов предложил (1954) модель триплетного Г. к., т. е. такого, в котором 1 аминокислоту кодирует группа из трёх нуклеотидов, наз. кодоном. Число возможных триплетов равно 43=64, а это более чем втрое превышает число распространённых аминокислот, в связи с чем было высказано предположение, что каждой аминокислоте соответствует несколько кодонов (т. н. вырожденность кода). Было предложено много различных моделей Г. к., из которых серьёзного внимания заслуживали три модели (см. рис.): перекрывающийся код без запятых, неперекрывающийся код без запятых и код с запятыми. В 1961 Ф. Крик (Великобритания) с сотрудниками получил подтверждение гипотезы триплетного неперекрывающегося кода без запятых. Установлены следующие основные закономерности, касающиеся Г. к.: 1) между последовательностью нуклеотидов и кодируемой последовательностью аминокислот существует линейное соответствие (колинеарность Г. к.); 2) считывание Г. к. начинается с определённой точки; 3) считывание идёт в одном направлении в пределах одного гена; 4) код является неперекрывающимся; 5) при считывании не бывает промежутков (код без запятых); 6) Г. к., как правило, является вырожденным, т. е. 1 аминокислоту кодируют 2 и более триплетов-синонимов (вырожденность Г. к. уменьшает вероятность того, что мутационная замена основания в триплете приведёт к ошибке); 7) кодовое число равно трём; 8) код в живой природе универсален (за некоторыми исключениями). Универсальность Г. к. подтверждается экспериментами по синтезу белка in vitro. Если в бесклеточную систему, полученную из одного организма (например, кишечной палочки), добавить нуклеиновокислотную матрицу, полученную из др. организма, далеко отстоящего от первого в эволюционном отношении (например, проростков гороха), то в такой системе, будет идти белковый синтез. Благодаря работам американских генетиков М. Ниренберга, С. Очоа, Х. Корана известен не только состав, но и порядок нуклеотидов во всех кодонах (см. табл., построенную по данным опытов с кишечной палочкой).
Из 64 кодонов у бактерий и фагов 3 кодона — УАА, УАГ и УГА — не кодируют аминокислот; они служат сигналом к освобождению полипептидной цепи с рибосомы, т. е. сигнализируют о завершении синтеза полипептида. Их назыают терминирующими кодонами. Существуют также 3 сигнала о начале синтеза — это т. н. инициирующие кодоны — АУГ, ГУГ и УУГ, — которые, будучи включенными в начале соответствующей информационной РНК (и-РНК), определяют включение формилметионина в первое положение синтезируемой полипептидной цепи. Приведённые данные справедливы для бактериальных систем; для высших организмов многое ещё не ясно. Так, кодон УГА у высших организмов может быть значащим; не совсем понятен также механизм инициации полипептида.
Реализация Г. к. в клетке происходит в два этапа. Первый из них протекает в ядре; он носит название транскрипции и заключается в синтезе молекул и-РНК на соответствующих участках ДНК. При этом последовательность нуклеотидов ДНК «переписывается» в нуклеотидную последовательность РНК. Второй этап — трансляция — протекает в цитоплазме, на рибосомах; при этом последовательность нуклеотидов и-РНК переводится в последовательность аминокислот в белке: этот этап протекает при участии транспортной РНК (т-РНК) и соответствующих ферментов (см. Белки, раздел Биосинтез).
Лит.: Общая природа генетического кода для белков, в сборнике: Молекулярная генетика, пер. с англ., М., 1963; Крик Ф., Генетический код (I), в кн.: Структура и функция клетки, пер. с англ., М., 1964, с. 9—23; Ниренберг М., Генетический код (II), там же, с. 24—41; Хэйс У., Генетика бактерий и бактериофагов, пер. с англ., М., 1965; Хартман Ф., Саскайнд З., Действие гена, пер. с англ., М., 1966; Бреслер С. Е., Введение в молекулярную биологию, 2 изд., М. — Л., 1966; Ингрэм В., Биосинтез макромолекул, пер. с англ., М., 1966; Лобашев М. Е., Генетика, 2 изд., Л., 1967; Уотсон Дж., Молекулярная биология гена, пер. с англ., М., 1967; Сойфер В. Н., Молекулярные механизмы мутагенеза, М., 1969; Дубинин Н. П.. Общая генетика, М., 1970.
Лингвистико-Волновой геном. Следующий шаг. №1
29 Окт 2010 18:12 #336
ППГ написал(а):
Даже простейший пример не хотят видеть
Селеноцистеин (selenocysteine) — нестандартная «двадцать первая» аминокислота, отличающаяся от обычного цистеина тем, что вместо атома серы в ее состав входит атом селена. Селеноцистеин является обязательным компонентом нескольких важных ферментов в организме животных (включая человека), простейших, бактерий и архей. Белки, содержащие селеноцистеин, называют селенопротеинами (selenoproteins). В геноме человека содержится более 20 генов селенопротеинов, поэтому селен является необходимым компонентом питания, и его недостаток в пище приводит к различным заболеваниям.
Стандартный генетический код, однако, предусматривает возможность кодирования только двадцати «канонических» аминокислот, и для того, чтобы закодировать двадцать первую, эволюции пришлось пойти на хитрость. Для кодирования селеноцистеина используется кодон UGA, который по умолчанию является стоп-кодоном, то есть сигнализирует об окончании синтеза белковой молекулы. Однако если за кодирующим участком гена следует особая последовательность нуклеотидов, которая называется SECIS (selenocysteine insertion sequence), аппарат синтеза белка интерпретирует кодон UGA как кодирующий селеноцистеин.
У организмов, в геноме которых есть гены селенопротеинов, имеется специальная селенопротеиновая транспортная РНК, которая распознает кодон UGA при помощи комплементарного ему антикодона UCA. Последовательность SECIS в ходе транскрипции включается в состав матричной РНК, синтезируемой на основе селенопротеинового гена. Оказавшись в составе матричной РНК, SECIS сворачивается в особую трехмерную структуру — «шпильку» с двумя петельками (см. рисунок), которая и служит сигналом, заставляющим аппарат синтеза белка присоединить к кодону UGA транспортную РНК, несущую селеноцистеин.
Антон Туранов и Вадим Гладышев из Университета штата Небраска (Линкольн, США) и их коллеги из нескольких американских научных институтов изучили механизм кодирования селеноцистеина у инфузории Euplotes. Эта инфузория интересна тем, что у нее, в отличие от других инфузорий и подавляющего большинства живых существ, кодон UGA не является стоп-кодоном и «по умолчанию» кодирует цистеин. Исследователи подумали, что если у Euplotes есть селенопротеины, то эта инфузория может оказаться единственным организмом, у которого один и тот же триплет нуклеотидов кодирует, в зависимости от контекста, две разные аминокислоты.
...
Таким образом, генетический код, «в норме» используемый для кодирования 20 аминокислот, при необходимости может быть расширен. Кроме селеноцистеина, который считают 21-й аминокислотой, существует и 22-я — пирролизин (pyrrolysine). Он входит в состав некоторых важных ферментов у организмов, считающихся одними из древнейших на планете, — архей-метаногенов (см. метаногенез). Пирролизин, как и селеноцистеин, кодируется каноническим стоп-кодоном, но другим (UAG), и тоже в сочетании со специальной регуляторной «шпилькой» в некодирующей области гена.
Лингвистико-Волновой геном. Следующий шаг. №1
29 Окт 2010 18:31 #338
Как известно, ДНК человека представляет собой «текст», составленный из миллиардов выстроенных в разных сочетаниях четырех «букв»-нуклеотидов - азотистых оснований аденина, гуанина, тимина и цитозина.
Музыканту и ученому доктору Эндрю Морли пришла в голову идея сопоставить с каждым из четырех нуклеотидов определенную ноту и получить так музыку генома.
Полученное таким образом музыкальное произведение, получившее название «Аллель» (генетический термин, обозначающий различные формы одного и того же гена), будет исполнено во вторник 13 июля Новым лондонским камерным хором в здании Королевского медицинского общества.
В этом произведении прозвучат генетические коды каждого из 40 участников хора.
В создании «геномного хорала» Морли помогал вдохновленный его идеей композитор Майкл Зев Гордон. «Я рассматривал генетический код с двух точек зрения: как сырой материал, который надо перевести на язык нот, а также как нечто чудесное, обладающее необыкновенной красотой. Так и выросло это произведение», - сказал Гордон.
Произведение начинается с простой ритмической фразы, которую поет один участник хора. Затем, по мере развития музыкальной темы, к нему присоединяются другие голоса, передавая биологический процесс репликации ДНК.
В ключевой момент произведения каждый из певцов начинает исполнять мелодию своего собственного генетического кода.
Лингвистико-Волновой геном. Следующий шаг. №1
29 Окт 2010 18:31 #339
С синонимами все ясно, омонимы НЕ ПОНЯТЫ. Упорно не понимаются. А поймут - модель летит... А рибосомы продолжают спокойно работать, читая синонимы и соображая какой смысл дать омонимам с учетом контекста. Вот он второй генетический код по Овчинникову (правда, он постеснялся обидеть папу-Крика) nature.web.ru/db/msg.html?mid=1157645 и не назвал вещи своими именами - МОДЕЛЬ КОДА НЕ ПОЛНА.
Лингвистико-Волновой геном. Следующий шаг. №1
29 Окт 2010 18:34 #340
ППГ написал(а):
Вот он второй генетический код
Старье, да старье - пора ППГ за школьную парту садиться:
[Третий генетический код, безусловно, существует, хотя его расшифровка еще до конца не закончена. Можно утверждать, что в отличие от универсального первого и второго генетических кодов третий код, работающий на стадии терминации трансляции, различен у эвкариот и прокариот, что не было предсказано ранее. Очевидно, что третий код не похож на второй, поскольку здесь взаимодействуют не две гомополимерные цепочки, как при кодон-антикодоновом взаимодействии, а линейная цепочка нуклеотидов (стоп-кодон) узнается разными фрагментами полипептидной цепи белкового фактора терминации класса-1 в трехмерном пространстве внутри рибосомы. Далее, скорее всего, точность декодирования третьего кода у эвкариот недостаточно высока, и поэтому возник дополнительный механизм проверки кодового соответствия, в котором, видимо, важную роль играет фактор класса-2 eRF3.
Лингвистико-Волновой геном. Следующий шаг. №1
29 Окт 2010 18:42 #341
Жаль, не могу привести мелодию ДНК по нашей технологии, когда музыка считывается не с табличного текста ДНК последовательностей, а прямо с препарата ДНК с генерацией мШЭИ ДНК с последующей комп. обработкой акустического варианта мШЭИ и извлечением мелодии ДНК как стратегической информации закона КРАСОТЫ И ГАРМОНИИ. Могу выслать в личку. Интересно, что файл такой музыки ДНК делится на музыку генов белков и РНК и музыку некодирующей ДНК, которая существенно отлична. Вероятно, голограммы (или то, что от них осталось в препаратах) не кодирующей, мусорной ДНК не поют, если они не прочитаны соответствующим образом.
Лингвистико-Волновой геном. Следующий шаг. №1
29 Окт 2010 23:39 #348
ППГ написал(а):
условие задачки - кодируют в триплетах только первые две буквы (Гамов был частично прав), третьи не участвуют в кодировании. Кодирование идет по схеме два из трех
Если два, то из четырёх, 4^2 = 16 и как закодировать ещё 4-5 аминокислот?
Лингвистико-Волновой геном. Следующий шаг. №1
30 Окт 2010 11:30 #352
Хайдук написал(а):
К чему удивляццо, Петр, ведь бывают и другие триплеты, что кодируют больше, чем одну аминокислоту?
'Удивляццо' на вас уже не буду, ваши проблемы. Напомню говоренное сотню раз и видимое здравым смыслом и экспериментом, что 32 ДУБЛЕТА-ОМОНИМА кодируют больше, чем одну аминокислоту (и Стоп-команду). Как выбрать точное значение кодона (дублета в нем)? Это логико-семантическая задачка, которую геном РЕШАЕТ, будучи соображающим (в отличие от хайдуков-генетиков). Это стратегическое положение. Неприятие его, игнорирование = ГМ еда = вырождение людей.
Лингвистико-Волновой геном. Следующий шаг. №1
30 Окт 2010 12:15 #354
ППГ написал(а):
Это вам ясно, а вот генетикам и молбиологам (части) - нет, мозги засохли
Это волногощикам засохли, а вот генетикам и молбиологам - нет:
The genetic code consists of 64 triplets of nucleotides. These triplets are called codons.With three exceptions, each codon encodes for one of the 20 amino acids used in the synthesis of proteins. That produces some redundancy in the code: most of the amino acids being encoded by more than one codon.
One codon, AUG serves two related functions:
* it signals the start of translation
* it codes for the incorporation of the amino acid methionine (Met) into the growing polypeptide chain
...
Codon Bias: All but two of the amino acids (Met and Trp) can be encoded by from 2 to 6 different codons. However, the genome of most organisms reveals that certain codons are preferred over others. In humans, for example, alanine is encoded by GCC four times as often as by GCG. This probably reflects a greater translation efficiency by the translation apparatus (e.g., ribosomes) for certain codons over their synonyms.
Exceptions to the Code: The genetic code is almost universal. The same codons are assigned to the same amino acids and to the same START and STOP signals in the vast majority of genes in animals, plants, and microorganisms. However, some exceptions have been found. Most of these involve assigning one or two of the three STOP codons to an amino acid instead.
Mitochondrial genes: When mitochondrial mRNA from animals or microorganisms (but not from plants) is placed in a test tube with the cytosolic protein-synthesizing machinery (amino acids, enzymes, tRNAs, ribosomes) it fails to be translated into a protein.
The reason: these mitochondria use UGA to encode tryptophan (Trp) rather than as a chain terminator. When translated by cytosolic machinery, synthesis stops where Trp should have been inserted.
In addition, most
* animal mitochondria use AUA for methionine not isoleucine and
* all vertebrate mitochondria use AGA and AGG as chain terminators.
* Yeast mitochondria assign all codons beginning with CU to threonine instead of leucine (which is still encoded by UUA and UUG as it is in cytosolic mRNA).
...
Nuclear genes: Violations of the universal code are far rarer for nuclear genes.
A few unicellular eukaryotes have been found that use one or two (of their three) STOP codons for amino acids instead.
Nonstandard Amino Acids: The vast majority of proteins are assembled from the 20 amino acids listed above even though some of these may be chemically altered, e.g. by phosphorylation, at a later time.
However, two cases have been found where an amino acid that is not one of the standard 20 is inserted by a tRNA into the growing polypeptide.
* selenocysteine. This amino acid is encoded by UGA. UGA is still used as a chain terminator, but the translation machinery is able to discriminate when a UGA codon should be used for selenocysteine rather than STOP. This codon usage has been found in certain Archaea, eubacteria, and animals (humans synthesize 25 different proteins containing selenium).
* pyrrolysine. In several species of Archaea and bacteria, this amino acid is encoded by UAG. How the translation machinery knows when it encounters UAG whether to insert a tRNA with pyrrolysine or to stop translation is not yet known. ]
Лингвистико-Волновой геном. Следующий шаг. №1
30 Окт 2010 12:33 #355
ППГ написал(а):
что 32 ДУБЛЕТА-ОМОНИМА кодируют больше, чем одну аминокислоту (и Стоп-команду). Как выбрать точное значение кодона (дублета в нем)? Это логико-семантическая задачка, которую геном РЕШАЕТ
Лингвистико-Волновой геном. Следующий шаг. №1
30 Окт 2010 12:38 #356
ППГ написал(а):
32 ДУБЛЕТА-ОМОНИМА кодируют больше, чем одну аминокислоту (и Стоп-команду). Как выбрать точное значение кодона (дублета в нем)?
Где видите эти 32 дублета, Петр? Дублетов из 4 нуклеотодов/букв 16 = 4^2. Чтобы закодировать оставшиеся 4 аминокислоты, ДНК не смог бы заставить даже ... ЛВГ
Лингвистико-Волновой геном. Следующий шаг. №1
30 Окт 2010 12:46 #359
Хайдук написал(а):
А Вы как его понимаете?
ППГ написал(а):
В традиционной триплетной модели генетического кода легко обнаруживается омонимии кодирующих дублетов. Значение таких омонимий до сих пор не понято и не оценено
P.S. Еще более не понятен смысл и значение омонимии кодирующих моноплетов...