Отлично. При малейшем проявлении хамства и неуважения ко мне - прекращаю. Все негативные эмоции внутри, от них никуда. Внешне поддерживаем вежливый разговор. Только логика и по возможности факты. Тон осуждения и обличения - в сторону. Это контр продуктивно, как много раз говорил. Ложь вообще недопустима. Ошибиться можно. Это обычное дело. И без кличек.
Итак начнем с чистого листа. Какой эксперимент побудил вас создавать ВГ Петрович? Есть ли экспериментальные данные, которые выявляли бы ошибки в предсказаниях модели Крика?
Вот например подробнейший обзор об измерениях точности трансляции на момент 1989 года. В нем по какой то причине проблемы с предсказаниями модели Крика не упоминаются. www.ncbi.nlm.nih.gov/pmc/articles/PMC372...crorev00042-0009.pdf
Итак ключевой вопрос, какой экспериментальный факт свидетельствует о необходимости пересмотра модели Крика?
Итак начнем с чистого листа. Какой эксперимент побудил вас создавать ВГ Петрович? Есть ли экспериментальные данные, которые выявляли бы ошибки в предсказаниях модели Крика?
Вот например подробнейший обзор об измерениях точности трансляции на момент 1989 года. В нем по какой то причине проблемы с предсказаниями модели Крика не упоминаются. www.ncbi.nlm.nih.gov/pmc/articles/PMC372...crorev00042-0009.pdf
Итак ключевой вопрос, какой экспериментальный факт свидетельствует о необходимости пересмотра модели Крика?
А давайте начнем с конца, где точки над i расставлены. Начнем, и может, сразу закончим дискуссию на том, что Ф.Крик, которого с десяток раз цитировал, в конце жизни в своей автобиографической книге "Безумный поиск" признал, что ИХ модель белкового кода "не имеет очевидного смысла". Очень умно сказал, что модель не вообще бессмысленная, но, мол, смысл есть, но НЕ ОЧЕВИДНЫЙ. Что он имел в виду сейчас уже не узнать. Хотелось бы думать, что он все-таки видел странность в не синонимических кодонах с их как бы очевидным двойным кодированием, которого не должно было быть по их с Ниренбергом представлениям. Его, двойного, и нет НА ВЫХОДЕ синтеза белков, но воблирование 3-го нуклеотида как бы обязывает к неопределенному двойному кодированию. Может, он и видел выход из противоречия их модели, но такой выход привел бы к вещам, к которым он не был готов. А именно, к признанию реальной текстовости белковых генов. Т.е. к мысли о разумности генетического аппарата, способного к контекстным смысловым ориентациям в многомерном смысловом пространстве мРНК. Все это увидел известный мол. биолог Ульф Лагеквист и дал даже формулу чтения кодона антикодоном "Два из трех", что автоматически приводит к ошибкам при чтении мРНК и синтезам ошибочных белков. Но Лагерквист пошел на сделку с совестью, сказав в своей статье, что такие кодоны с двойным кодированием встречаются крайне редко, что неверно. Даже если таких двойственно кодирующих триплетов всего один или два (например, кодон UUU), то этого достаточно, чтобы аномально исказить создаваемый белковый пейзаж. Итак, проблема подвисла без решения.
Теперь ответ на ваш конкретный вопрос относительно предсказаний ошибок модели. Тут удар модели нанесен практикой: это перекодировки кодонов в стрессовых условиях - аминокислотное голодание, тепловой шок, наличие антибиотиков в клетке. По Крику - кодоны строго однозначны и никаких перекодировок быть не должно. А они есть. Второе доказательство неадекватности модели Крика-Ниренберга - это скандал вокруг статьи Lolle, Pruitt et al в Nature 2005г. о работе гена Ноt Head в растении A.thaliana. Но это отдельный разговор.
PP: "Тогда, когда вы начнете аргументировать свои утверждения. Например обоснуете чем вас не устраивает масс спек, который выдает один пик и как раз в том месте, где и положено для продукта с учетом его массы".
ППГ: Насколько помню, и указывал на это, приведенные масс спектры пептидов (а я занимался масс спектрометрией) давали НЕ ОДИН пик молекулярных ионов пептидов, но уширенные пики за счет наличия близких, но не одинаковых по массе пептидов. Это косвенно свидетельствует о вариациях аминокислот в синтезируемых пептидах, что и дает пептидам разброс по массам. Такие вариации могут быть следствием контекстных игр на мРНК с включением разных аминокислот в пептиды, хотя они могут кодироваться одними и тем же не синонимическими кодонами-омонимами с переменчивым кодированием, зависящим от контекста мРНК.
Дык когда мы услышим хоть, что то похожее на ответ связанный с вопросом..
Есть ли экспериментальные данные, которые выявляли бы ошибки в предсказаниях модели Крика?
Кроме общих монологов о том, что кто то и где то чем то пытался заниматься ..
ps Медведь во мне проснулся, не иначе,
Чувствую, как медленно впадаю в спячку.
странность в не синонимических кодонах с их как бы очевидным двойным кодированием, которого не должно было быть по их с Ниренбергом представлениям. Его, двойного, и нет НА ВЫХОДЕ синтеза белков, но воблирование 3-го нуклеотида как бы обязывает к неопределенному двойному кодированию.
кто такое двойное кодирование, Петрович?
nonlocality wrote:
кодоны с двойным кодированием встречаются крайне редко, что неверно. Даже если таких двойственно кодирующих триплетов всего один или два (например, кодон UUU), то этого достаточно, чтобы аномально исказить создаваемый белковый пейзаж.
1-2 из 64 это действительно редко и не знаю как может исказить белковый пейзаж
nonlocality wrote:
перекодировки кодонов в стрессовых условиях - аминокислотное голодание, тепловой шок, наличие антибиотиков в клетке. По Крику - кодоны строго однозначны и никаких перекодировок быть не должно. А они есть.
раз есть, то где и в чём состоят эти конкретные примеры?
А давайте начнем с конца, где точки над i расставлены. Начнем, и может, сразу закончим дискуссию на том, что Ф.Крик, которого с десяток раз цитировал, в конце жизни в своей автобиографической книге "Безумный поиск" признал, что ИХ модель белкового кода "не имеет очевидного смысла". Очень умно сказал, что модель не вообще бессмысленная, но, мол, смысл есть, но НЕ ОЧЕВИДНЫЙ.
Нет давайте начнем не с конца, а с начала. Автобиография не есть научный труд. Смысл может и не очевидный, но модель то работает. Эйнштейну например был не очевиден смысл квантовой механики, это же не повод от нее отказываться.nonlocality wrote:
Теперь ответ на ваш конкретный вопрос относительно предсказаний ошибок модели. Тут удар модели нанесен практикой: это перекодировки кодонов в стрессовых условиях - аминокислотное голодание, тепловой шок, наличие антибиотиков в клетке. По Крику - кодоны строго однозначны и никаких перекодировок быть не должно. А они есть. Второе доказательство неадекватности модели Крика-Ниренберга - это скандал вокруг статьи Lolle, Pruitt et al в Nature 2005г. о работе гена Ноt Head в растении A.thaliana. Но это отдельный разговор.
Вы же ВГ создали задолго до 2005 года. Вот я интересуюсь, какой эксперимент/явление побудил вас в те годы это делать. Обычно от старых теорий отказываются, когда обнаруживают границы их применимости, за которыми они не работают, и требуется идти дальше. Так вот что побудило? Кстати когда вы говорите о перекодировке кодонов, вы дайте пожалуйста ссылку, чтобы было ясно о чем вы.
nonlocality wrote:
перекодировки кодонов в стрессовых условиях - аминокислотное голодание, тепловой шок, наличие антибиотиков в клетке. По Крику - кодоны строго однозначны и никаких перекодировок быть не должно. А они есть.
раз есть, то где и в чём состоят эти конкретные примеры?
Quote Selected
«Во многих работах показано, что ошибочная трансляции белков рибосомой вызывается разнообразными неблагоприятными факторами антибиотиками, изменением температуры, созданием определенных концентраций катионов, аминокислотным голоданием и другими условиями внешней среды. Повышенная неоднозначность трансляции кодонов, локализованных в особом контексте, имеет биологическое значение и приводит к неслучайному распределению “ошибочных” аминокислот по длине синтезируемого полипептида, приводящему к модификациям функций белков с выходом на механизмы клеточных дифференцировок, и поэтому контексты иРНК являются субстратом естественного отбора. Оптимальный уровень “ошибок” трансляции (если это действительно ошибки) регулируется неизвестными механизмами, и он онтогенетически и эволюционно оправдан».
Тер-Аванесян М.Д., Инге-Вечтомов С.Г. Генетический контроль синтеза белка.. Изд. Ленинградского Университета .Л., 1988. С.29
Нет давайте начнем не с конца, а с начала. Автобиография не есть научный труд. Смысл может и не очевидный, но модель то работает. Эйнштейну например был не очевиден смысл квантовой механики, это же не повод от нее отказываться.nonlocality wrote:
Не модель работает, но сам код. PP wrote:
Вы же ВГ создали задолго до 2005 года. Вот я интересуюсь, какой эксперимент/явление побудил вас в те годы это делать. Обычно от старых теорий отказываются, когда обнаруживают границы их применимости, за которыми они не работают, и требуется идти дальше. Так вот что побудило? Кстати когда вы говорите о перекодировке кодонов, вы дайте пожалуйста ссылку, чтобы было ясно о чем вы.
Еще на 5 курсе биофака МГУ как-то повнимательнее посмотрел канон-таблицу кода и вдруг увидел ее, НУ НЕ ТО ЧТОБЫ ИДИОТИЗМ, НО ЧТО-ТО ВРОДЕ. Не синонимы почему-то делали двойное кодирование, несовместимое с моделью Крика-Ниренберга. Вот и вся история.
Еще на 5 курсе биофака МГУ как-то повнимательнее посмотрел канон-таблицу кода и вдруг увидел ее, НУ НЕ ТО ЧТОБЫ ИДИОТИЗМ, НО ЧТО-ТО ВРОДЕ. Не синонимы почему-то делали двойное кодирование, несовместимое с моделью Крика-Ниренберга. Вот и вся история.
Ну это чисто умозрительные соображения. Опять возьмем квантовую механику как пример. Электрон летит сразу через две щели это же тоже можно назвать идиотизмом. Но теория отлично работает. А вы получается начали строить новую теорию на пустом месте. ВГ получается не была созадана чтобы объяснить конкретный феномен, а уже постфактум вы пытались подогнать под нее что либо, даже такие явления, которых и нет в природе, типа телегонии. Вы не находите, что вы таким образом отступили от научного метода? nonlocality wrote:
Во многих работах показано, что ошибочная трансляции белков рибосомой вызывается разнообразными неблагоприятными факторами антибиотиками, изменением температуры, созданием определенных концентраций катионов, аминокислотным голоданием
Ну это как бы очевидный факт. Скорость химических реакций зависит от концентраций аминокислот, изменения внешних факторов очевидно будут влиять константы по разному для разных аминокислот. Поэтому уровень ошибок, не такой уж маленький изначально (1:1000), будет очевидно меняться. Никаких противоречий с моделью Крика тут очевидно нет.
Ну это как бы очевидный факт. Скорость химических реакций зависит от концентраций аминокислот, изменения внешних факторов очевидно будут влиять константы по разному для разных аминокислот. Поэтому уровень ошибок, не такой уж маленький изначально (1:1000), будет очевидно меняться. Никаких противоречий с моделью Крика тут очевидно нет.
Ну это чисто умозрительные соображения. Опять возьмем квантовую механику как пример. Электрон летит сразу через две щели это же тоже можно назвать идиотизмом. Но теория отлично работает. А вы получается начали строить новую теорию на пустом месте. ВГ получается не была созадана чтобы объяснить конкретный феномен, а уже постфактум вы пытались подогнать под нее что либо, даже такие явления, которых и нет в природе, типа телегонии. Вы не находите, что вы таким образом отступили от научного метода?
ППГ: М.Ниренберг и Ф.Крик в соей статье 1964г. в УФН, которую не раз цитировал (и Вика тоже) указали на собственные эксперименты, где поликодон UUU cтимулировал одновремнное включение в синтезируемый пептид фенилаланина и лейцина, что исключало их постулат об однозначном кодировании. Модель уже от этого факта становилась ложной. Позже появилась статья, где экспериментально доказано, что один из кодонов омонимов одновременно кодирует аминокислоту и стоп. Эту статью здесь привёл, насколько помню, Олег. И наконец, если модель демонстрирует явное и видимое всем противоречие, то логично его обнародовать, что и сделал У.Лагерквист, но испугался авторитетов двух нобелиатов и смазал концовку своей статьи, которую тоже не раз цитировал. И последнее - нет проверок коллинеарностей аминокислот и кодонов, кроме одного случая с серповидной анемией. И то по одному кодону. Упоминал о эксп. работе Лолли, Прюита и соавт. 2005г. в Nature. У них результаты противоречили законам Менделя. Мутантный ген Hot Head, отвечающий за аномалию в фенотипе A.thaliana (мутантная замена кодона) в определенном проценте проросших семян переставал давать аномальный фенотипический ответ, и растения возвращались к дикому типу. По классической генетике такого в принципе не может быть. Но было. Понять этого никто не мог. Был скандал. Авторов обвинили во всех мыслимых и не мыслимых грехах. Лолли потеряла работу. А объяснение простое - в контекстных перестройках ДНК вокруг мутантного гена: кодовое значение аномального кодона вернулось к прежнему исходному значению - и фентоип вернулся к дикому типу. Мы написали письмо Прюиту, но тот был, видимо, слишком напуган скандалом, а тут еще про перекодировки ему толкуем. Не ответил.
nonlocality wrote:
Во многих работах показано, что ошибочная трансляции белков рибосомой вызывается разнообразными неблагоприятными факторами антибиотиками, изменением температуры, созданием определенных концентраций катионов, аминокислотным голоданием
PP wrote:
Ну это как бы очевидный факт. Скорость химических реакций зависит от концентраций аминокислот, изменения внешних факторов очевидно будут влиять константы по разному для разных аминокислот. Поэтому уровень ошибок, не такой уж маленький изначально (1:1000), будет очевидно меняться. Никаких противоречий с моделью Крика тут очевидно нет.
ППГ: Кем это было доказано? Ссылку, пожалуйста, на "влиять константы". Это будет научный метод доказателств, а не вольные фантазии.
ППГ: Насколько помню, и указывал на это, приведенные масс спектры пептидов (а я занимался масс спектрометрией) давали НЕ ОДИН пик молекулярных ионов пептидов, но уширенные пики за счет наличия близких, но не одинаковых по массе пептидов. Это косвенно свидетельствует о вариациях аминокислот в синтезируемых пептидах, что и дает пептидам разброс по массам.
В статье четко указывается разрешающая способность метода. Авторы могут различить в виде пика замену одной аминокислоты, что они и делают
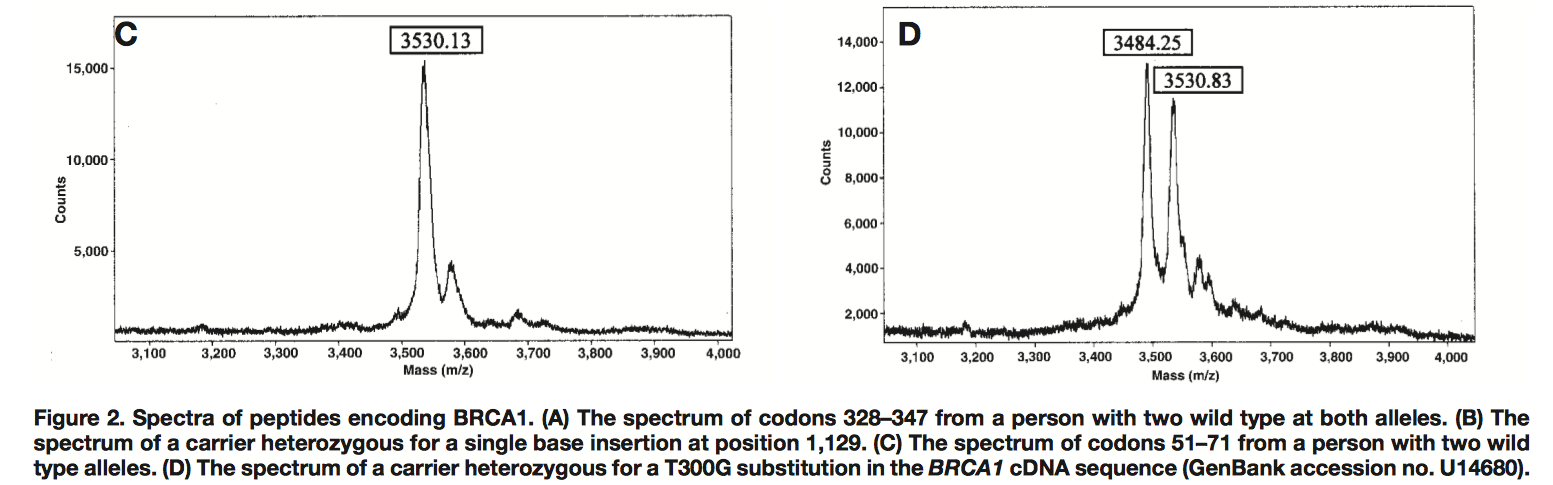
Слева wild type, справа гетерозиготная мутация в результате которой наблюдаем второй пик (3484.25). ВГ действительно предсказывает отсутствие пиков, ибо количество равновероятных омонимичных продуктов разной массы комбинаторно высоко, но как мы легко видим эксперимент это предсказание опровергает.
wild type: CCT TCA CAG TGT CCT TTA TGT -> mutant: CCT TCA CAG GGT CCT TTA TGT
Как видим омонимы есть (e.g. CA, TG, TT ), а пиков им соответствующих нет. Вот так несмотря на полное отсутствие контекста (транслируем всего 7 кодонов) трансляция идет точно как учит Крик.
ППГ: Кем это было доказано? Ссылку, пожалуйста, на "влиять константы". Это будет научный метод доказателств, а не вольные фантазии.
Я хочу уточнить, что вы называете вольными фантазиями, что скорость химических реакций зависит от концентраций, и что константы зависят от внешних факторов? Ну это даже в школьном учебнике химии должно быть прописано. Читайте любой учебник по физической химии если не верите.
P.S.
Перед тем как отсылать оппонента вам что либо доказывать, вы как автор революционной теории, обязаны убедить оппонентов, что вот такие обычные физико химические причины не могут объяснить наблюдаемых изменений в точности трансляции. Потом вы должны показать, что ВГ дает более точное объяснение точности трансляции в условиях стресса. А потом уже оппоненты будут обязаны реагировать.
К сожалению становится видно, что построения вИдения процессов связанные с ЭМ-полями у ППГ от таблицы “неправильности чтения” кодонной решетки данных несколько преувеличено. Приходится согласится, что основным внешним фактором становится следующий постулат сказанный пользователем РР..
….Скорость химических реакций зависит от концентраций аминокислот, изменения внешних факторов очевидно будут влиять константы по разному для разных аминокислот. ..
И это является еще раз дополнением, к тому. что ЭМ-рой подобных концентраций может быть управляем, имеет возможность внешнего перестроения и естественно может оказывать влияние на выбор “мишени” для действия. Естественно есть и свои ограничения. В этом случае основные требования , по той же трансляции, будут соблюдены, но вносится новая реальность - внешнее управление концентрациями и перестановками..
ps - Кстати, Семён Маркович, давно хотел вам сказать: то, что вы доктор
физико-математических наук, ещё не даёт вам право угрожать людям
заточкой.
Как видим омонимы есть (e.g. CA, TG, TT ), а пиков им соответствующих нет. Вот так несмотря на полное отсутствие контекста (транслируем всего 7 кодонов) трансляция идет точно как учит Крик.
Надо мне подробнее ознакомиться со статьей. Но пока основное возражение то же - мы видим размытые пики молекулярных ионов, что говорит о широком распределении масс синтезируемых пептидов, часть из которых может быть результатом контекстных манипуляций. С другой стороны, используемая 7-членная организация кодонов может быть недостаточной для определения смыслов их омонимов, поэтому выбор включаемых в пептид аминокислот будет случайным. Пример для пояснения. Фраза: "красивая коса". Контекст для слова "коса" короток и точно дешифровать омоним "коса" невозможно, выбор смыслов будет случайным. Но фраза "красивая девичья коса" имеет более длинный контекст и смысл омонима будет точным. Поэтому, экстраполируя это на синтез пептидов, мы в праве ожидать широкий набор трактовок кодонов-омонимов и, как следствие, синтез набора близких по массе пептидов, что и наблюдается в эксперименте. Этим нюансам Ф.Крик не учит. Сами должны соображать. Ф.Крик чувствовал слабость их модели, поэтому и произнес "смысл модели кода не очевиден". Очевидность смысла кодонов-омонимов достигается ПОЛНЫМ контекстом мРНК. Мера полноты контекстов пока не ясна. Равно как и смыслы мРНК и квази сознания генома. Это работа на весь 21 век.
Я хочу уточнить, что вы называете вольными фантазиями, что скорость химических реакций зависит от концентраций, и что константы зависят от внешних факторов? Ну это даже в школьном учебнике химии должно быть прописано. Читайте любой учебник по физической химии если не верите.
Из контекста нашего разговора ясно, что я высказываю сомнение именно к обсуждаемой ситуации, а не вообще сомневаюсь о роли скоростей хим. реакций. Отсылка к учебникам является нечестным передергиванием смыслов.
Кстати, перекодировки значений омонимических кодонов, зависящие от экстремальных флуктуаций внешней среды, как раз подтверждает мои соображения, что кодоны омонимы есть барометр неблагоприятных условий окружающей биосистему среды. Они обеспечивают синтез пробных белков с измененным аминокислотным составом. В пределе - это синтез иммуноглобулинов по Ву-Кэботу.
На пост nonlocality
К сожалению, нет. Ваши высказывания, лишены просто фактов. Одни так сказать “смыслы” которые не имеют никакого практического применения, так как оторваны от реальных событий и данных. Вами совершена ошибка (примерно как бы ошибка “второго рода”), когда Вы перескакиваете с понятий реальных химических и электрохимических взаимодействий на словесную пургу - заменяя их как бы “словосочетанями” или “смыслами”. И далее играете этими словосочетаниями отдаляя практику от виртуальной гипотезы. По мере Вашего разоблачения этих “видений” Вы, как обычно дополняете эту “пургу” - добавляя новые требования “смыслы” типа
Очевидность смысла кодонов-омонимов достигается ПОЛНЫМ контекстом мРНК. Мера полноты контекстов пока не ясна. Равно как и смыслы мРНК и квази сознания генома. Это работа на весь 21 век.
ps Новости из мира инноваций. По данным рейтингового агентства «Таймс-РФ» самым высокоэффективным коммерческим продуктом российской науки являются платные защиты кандидатских и докторских диссертаций.
Не вдаваясь в логику Гаряева, на данный момент пересмотра модели триплепного кода никто не требует.
Особенно те, кто синтезирует геномы.
Примеры перекодировок кодонов часто объясняются выбором аа-тРНК или мутациями в генах тРНК, как в случаях супрессии (неудачная ссылка на Вечтомова у него там), не нарушая триплетности.
=====
дискуссия на уровне -
15
ППГ-\\Во многих работах показано, что ошибочная трансляции белков рибосомой вызывается разнообразными неблагоприятными факторами антибиотиками, изменением температуры, созданием определенных концентраций катионов, аминокислотным голоданием\\
РР-\\Ну это как бы очевидный факт. Скорость химических реакций зависит от концентраций аминокислот, изменения внешних факторов очевидно будут влиять константы по разному для разных аминокислот. Поэтому уровень ошибок, не такой уж маленький изначально (1:1000), будет очевидно меняться. Никаких противоречий с моделью Крика тут очевидно нет. \\
Скорость химических реакций зависит от концентраций аминокислот, изменения внешних факторов очевидно будут влиять константы по разному для разных аминокислот. Поэтому
На пост nonlocality
К сожалению, нет. Ваши высказывания, лишены просто фактов. Одни так сказать “смыслы” которые не имеют никакого практического применения, так как оторваны от реальных событий и данных. Вами совершена ошибка (примерно как бы ошибка “второго рода”), когда Вы перескакиваете с понятий реальных химических и электрохимических взаимодействий на словесную пургу - заменяя их как бы “словосочетанями” или “смыслами”. И далее играете этими словосочетаниями отдаляя практику от виртуальной гипотезы. По мере Вашего разоблачения этих “видений” Вы, как обычно дополняете эту “пургу” - добавляя новые требования “смыслы” типа
Очевидность смысла кодонов-омонимов достигается ПОЛНЫМ контекстом мРНК. Мера полноты контекстов пока не ясна. Равно как и смыслы мРНК и квази сознания генома. Это работа на весь 21 век.
Давайте поосторожней в выражениях, иначе тот же путь в г.... Будьте благовоспитанным мальчиком.
Реальных событий у нас полно. Зря обольщаетесь. Обсуждать их здесь дело бесполезное, поэтому ограничусь пока теорией генетического кодирования, включающей такие понятия как 'смыслы мРНК' и 'квази сознание генома'. Вам пока это недоступно. Но ничего, постепенно поймете. Нет - ваши проблемы.