Ваша ошибка в том, что "Группа Чукч..." НЕ РАЗБИРАЕТ работу устройств для создания белков, они инициализировали расследования о том, КАК нужный материал попадает ИМЕННО в те места, где и происходит ЭТА сборка.
Именно пространственная цепочка по взаимодействию в пространстве наборов тРНК (со своими АК) и есть тот набор который "Группа Чукч..." выделила для этого.
Именно этот факт, что случайность этого события явно не соответствует, так все это происходит в нарушения случайного броуновского движения молекул в этой среде и говорит о том, что ВЗАИМОДЕЙСТВИЕ этих наборов тРНК для продвижения к месту сборки белков СУЩЕСТВУЕТ.
Вот и идет ощупывания этого процесса и его анализ, отчего и как...
Никакого "ощупывания" у чукч как не было, так и нет. Есть лженаучная профанация.
С другой стороны, есть идея голографического распознавания аминоацилированных комплексов (аа-тРНК) на основе геномной голографии Бегущей Волны интенсивности, данной А.А.Корнеевым в нашей статье (ссылку давал). После распознания по отдельности каждой из аа-тРНК, включая мРНК-контекстно выбранные аа-сиомы-тРНК, рибосомный клеточный нанобиокомпьютер принимает решение об упорядоченном в пространстве-времени включении каждой из распознанных аа-тРНК в растущую пептидную цепь, что и запускает биосинтез белков. Таким образом, ориентация аа-тРНК в пространстве-времени перед началом биосинтезов белков является ключевым фактором. Силовые процессы ориентации остаются пока неизвестными. Идет анализ и работа в этом направлении.
Но чукчи тут вне игры, поскольку тавот-аэродромная логика бражно-пекарско-дрожжевых процессов не приспособлена для такой работы.
О Блаженный Ботаник, опять запел свои голографические песни. Еще раз напоминаем, что тут разбирают вопрос как АК попадают в нужное место. Какие силы сдвигают эти цепочки из всего пространства клетки именно в место где собираются белки.
ps Шизофрения - странный предмет: ты вроде пингвин, а вроде омлет.
О Блаженный Ботаник, опять запел свои голографические песни. Еще раз напоминаем, что тут разбирают вопрос как АК попадают в нужное место. Какие силы сдвигают эти цепочки из всего пространства клетки именно в место где собираются белки.
Разве лженаучные безграмотные "суждения" НЕ СПЕЦИАЛИСТОВ в молекулярной биологии могут всерьёз восприниматься? Голографическая модель как раз объясняет пространственно-временную схему организации аа-тРНК ПЕРЕД началом биосинтезе белков. Это уже шаг вперед. Силовые, векторно двигающие процессы пока неясны. Хотя можно представить, что броунирующие аа-тРНК, находящиеся в цитозоле на ангстремных расстояниях канонических водородных связей (А-У, Г-Ц) уже будут в радиусе их воздействия и, соответственно, будут точно выстроены-селекционированы для сброса распознанных и нужных аминокислот в растущую пептидную цепь. Необходимы эксперименты, нацеленные на предлагаемую модель. И они будут.
Бредни чукч оставляем на их "научной" совести.
Блаженный Ботаник в своем амплуа - голографическая тень от работы Лазаря как двигатель прогресса. Единственно нужное из этого скрипа идей престарелого ученогоследующее..
...Силовые, векторно двигающие процессы пока неясны. Хотя можно представить, что броунирующие аа-тРНК, находящиеся в цитозоле на ангстремных расстояниях канонических водородных связей (А-У, Г-Ц) уже будут в радиусе их воздействия и, соответственно, будут точно выстроены-селекционированы для сброса распознанных и нужных аминокислот в растущую пептидную цепь...
В переводе на общепринятый язык - нет явных возражений, что молекулы тРНК и АК и их местонахождение в объеме клетки ОТЛИЧАЮТСЯ от случайного броуновского движения в этой структуреb и каким-то образом взаимодействуют. Это и есть базовое утверждение на котором может держаться дальнейшая дискуссия.
ps - И еще, согласно правилам нашей фирмы, вы никому не должны разглашать величину вашей заплаты.
- Да я собственно и не собирался позориться.
Источником излучения по гипотезы "Группы Чук..." это место "расплетения" на ДНК. В этом месте "расплетения" как раз и активизирован набор из 20 АК... из-за существования ЭМ-полей на самой ДНК. Это инициализатор ВСЕХ движений в клеточной структуре для синтеза белков для этого места. Рибосома это лишь один из участников этого процесса. мРНК это лишь один из первых этапов, получаемых из внешних АК и сложенных (информация) с места "расплетения" на ДНК.
каша у Кака полная: только нуклеотидов-кодонов может быть на месте "расплетения" на ДНК, никаких АК там нет; мРНК тоже из нуклеотидов ДНК получена, а не "из внешних АК".
даже если будет (а его нет) излучение в результаправит нужную тРНК (с АК) ИМЕННО в нужное место зборки, КАК выстроит цепоте "расплетения", то КАК оно начку тРНК с АК устремлённую ИМЕННО в нужное место зборки в рибосоме??
В переводе на общепринятый язык - нет явных возражений, что молекулы тРНК и АК и их местонахождение в объеме клетки ОТЛИЧАЮТСЯ от случайного броуновского движения в этой структуреb и каким-то образом взаимодействуют. Это и есть базовое утверждение на котором может держаться дальнейшая дискуссия.
Так это даже клопу ясно... А дальше надо соображать профессионально, но не по клопиному.
Ну вот хоть по одному из под вопросов пришли к согласию.
Теперь нужно убедить пользователя Хайдук (и других сомневающихся), что это и есть фрагмент варианта "ньютоновского" яблока на базе которого можно строить гипотезы, ну например, а ПОЧЕМУ идет это движение, например на базе тРНК (с АК) и ИМЕННО в нужное место для последующей сборки см.пп.11012, естественно внеся поправки (учитывая мнения пользователя Хайдук).
* в месте "расплетения" на ДНК АК входят в сборку нуклеотидов-кодонов вот они то и участвуют опосредовано в излучение ЭМ-поля по своим индивидуальным параметрам именно в этом месте.
* пока же идет разбор ПОЧЕМУ возможно и выстраиваются в пространстве молекулы тРНК (с АК) с помощью ЭМ-полей, возможно ли внешнее вмешательство в это построение или как.
ps Статистически даже самая умная голова обычно проигрывает в споре с тупым предметом.
Ну вот хоть по одному из под вопросов пришли к согласию.
Теперь нужно убедить пользователя Хайдук (и других сомневающихся), что это и есть фрагмент варианта "ньютоновского" яблока на базе которого можно строить гипотезы, ну например, а ПОЧЕМУ идет это движение, например на базе тРНК (с АК) и ИМЕННО в нужное место для последующей сборки см.пп.11012, естественно внеся поправки (учитывая мнения пользователя Хайдук).
* в месте "расплетения" на ДНК АК входят в сборку нуклеотидов-кодонов вот они то и участвуют опосредовано в излучение ЭМ-поля по своим индивидуальным параметрам именно в этом месте.
* пока же идет разбор ПОЧЕМУ возможно и выстраиваются в пространстве молекулы тРНК (с АК) с помощью ЭМ-полей, возможно ли внешнее вмешательство в это построение или как.
Тут можно много напридумывать. Самое простое, то, что уже сказал выше - случайное броунирование аа-тРНК мгновенно прекращается как только адекватный каноническим пАрам антикодон оказывается в поле водородных связей А-У, Г-Ц. И этот кодон выдергивается, а вслед за ним и нужная аминокислота.
Другое объяснение. Игра идет по спинтронной голограмме нуклеотидов в рамках голографии бегущей волны интенсивности по Корнееву. Тут проверить будет сложнее.
Ещё раз стоит напомнить П.Гаряеву, что разговор идёт о том как собираются тРНК в нужном месте. Дальнейшие действия ПОКА не рассматриваются. Чтобы это было выполнимо НУЖНО
1. Для этого нужно иметь некий центр дающий переодически ЭМ поле на определенных частотатах. Это центр для сбора.
2. Нужные молекулы должны иметь приемное устройство для принятия этого индивидуального частотного сигнала.
3. Эти молекулы должны транслировать этот сигнал далее, т.е. они должны быть осцилляторами.
4. Если эти условия выполняются и при каждой трансляции сигнал с не большим затуханием передаётся далее - можно говорить о создание цепочки взаимодействующих молекул в пространстве и возможности их поступательного сближения с заявленным центром сближения.
Ещё раз стоит напомнить П.Гаряеву, что разговор идёт о том как собираются тРНК в нужном месте. Дальнейшие действия ПОКА не рассматриваются.
А я напоминаю вам, что в варианте действия водородных связей А-У, Г-Ц по вытягиванию нужного кодона, не надо как-то особо собираться в нужном месте. Достаточно действия кодон-антикодоновых водородных связей на броунирующие в цитозоле аа-тРНК, связей, которые вытягивают нужный антикодон в составе аа-ТРНК. Это водород-связевое "сито", отбирающее нужные аа-РНК.
Водородная связь относится к понятию "слабая связь" радиус воздействия составляет ≈160 до 200 пм, что полностью делает не возможность вмешиваться в "дальнее" взаимодействие между молекулами тРНК в растворе
Водородная связь относится к понятию "слабая связь" радиус воздействия составляет ≈160 до 200 пм, что полностью делает не возможность вмешиваться в "дальнее" взаимодействие между молекулами тРНК в растворе
Опять клопиные гугловские "знания".
В варианте использования водородных связей, а оно имеет место для канон пар A-У, Г-Ц, о дальних взаимодействиях речи НЕ идет. Броунирование аа-тРНК в клеточном цитозоле обеспечивает попарные ближние взаимодействия их азотистых оснований, достаточные для избирательного вытягивания аа-тРНК по водородным связям их азот. оснований. Но это от вас отскакивает почему-то.
случайное броунирование аа-тРНК мгновенно прекращается как только адекватный каноническим пАрам антикодон оказывается в поле водородных связей А-У, Г-Ц. И этот кодон выдергивается, а вслед за ним и нужная аминокислота.
В варианте использования водородных связей, а оно имеет место для канон пар A-У, Г-Ц, о дальних взаимодействиях речи НЕ идет. Броунирование аа-тРНК в клеточном цитозоле обеспечивает попарные ближние взаимодействия их азотистых оснований, достаточные для избирательного вытягивания аа-тРНК по водородным связям их азот. оснований.
ну вот видите, Петрович, как всё просто и НЕТ надобности в ваших сиомах, голографии и прочей фигне
Опять "во городе бузина в Киеве дядька", т.е. П. Гаряев не понимает, что разговор идет о том, как и каким образом тРНК из окраины клетки попадает внутрь ядра и к месту "расплетения". Водородные связи для дрейфа тРНК от центра до самой до окраины на таком расстояние явно не сработают.
Предложенная схеме работы пользователем Поршень, как раз и укладывается в схему движения тРНК со своими особенностями. Причем тут не нужны ни вакуум по Шипову, ни миражи голограмм, ни темная материя.
Именно это и пугает нашего форумного ботаника.
ps Нас помнят, пока мы мешаем другим.
по-видимому, чукча, тРНК всяких мастей ошиваются вокруг да около рибосом (мест синтеза белков), а НЕ вокруг места "расплетения" ДНК - там им делать нечего, там лишь цепочка мРНК может выстраиваться, дабы потом подойти к рибосоме и начать выхватывать очередную тРНК с АК верхом из окружающего облака/супчика тРНК всяких мастей
Мы сейчас пока не трогаем эти тоже нужные вопросы. Дело в том, что все эти действия происходят вблизи "расплетения" на ДНК, так сказать в шаговой доступности. Пока реально обсуждается вопрос стимуляции подхода множественных цепочек однотипных тРНК из разных мест их зарождения, к этому нужному месту для сборки.
ps
- Привет, не спишь ещё?
- Нет.
- Ну я позвоню позже.
ну вот видите, Петрович, как всё просто и НЕТ надобности в ваших сиомах, голографии и прочей фигне
О сиомах в этой части рассуждений не пишу, чтобы не усложнять.
При "водородных выдергиваниях" сиомия и её проблема никуда не деваются. Решение их остаются те же, что вы не поняли, точнее, не прочитали. Для 3-й виртуально вобулирующей буквы кодона срабатывает принцип "делегирования" перекодировочного смысла для этой буквы. Как в примере: "Я забыл коГ своей квартиры и не могу войти". Контекст ситуации (читай мРНК, где данный сиом-кодон) диктует мне делегировать букве Г смысл буквы Д, т.е. перекодировать Г на Д. И задача решена - вы в собственной квартире. Видите, как просто. Надо только усвоить, что мРНК - это реальный текст с определенным смыслом, а не метафорическое тупое, по сути, сравнение последовательности нуклеотидов мРНК с текстом, как принято в мол. биологии. Долой метафоричность! Не будьте бабусей, гордящейся своим острым зрением и забывшей, что у нее на носу "контекст-очки".
Опять "во городе бузина в Киеве дядька", т.е. П. Гаряев не понимает, что разговор идет о том, как и каким образом тРНК из окраины клетки попадает внутрь ядра и к месту "расплетения". Водородные связи для дрейфа тРНК от центра до самой до окраины на таком расстояние явно не сработают.
Предложенная схеме работы пользователем Поршень, как раз и укладывается в схему движения тРНК со своими особенностями. Причем тут не нужны ни вакуум по Шипову, ни миражи голограмм, ни темная материя.
Именно это и пугает нашего форумного ботаника.
Это вы, любезный, воспитанный в тавот-дрожжевой атмосфере, никак не усвоите школьное (как я студентом ляпнул акад. РАН Сисакяну о долгом и трудном путешествии аа-тРНК к нужному кодону мРНК). В цитозоле (цитоплазме) клеток всегда в избытке пул разных аа-тРНК. И не надо никуда "попадать". Идет избирательное "выдергивание" нужных аа-тРНК на комплекс "мРНК-рибосома" по водородным связям. А темная материя, голография, вакуум - это уже другая история, не по отсутствующим зубам чукч.
Опять Блаженный Ботаник в своем амплуа. Поясняем этому старцу от науки, разговор идет НЕ о том, КАК к молекуле РНК(любого вида) присоединяются к ней АК а ОТКУДА они появляются в нужном для этого месте (что на ДНК для иРНК, что в другом месте для тРНК, да и как тРНК мигрируют в самой клете)
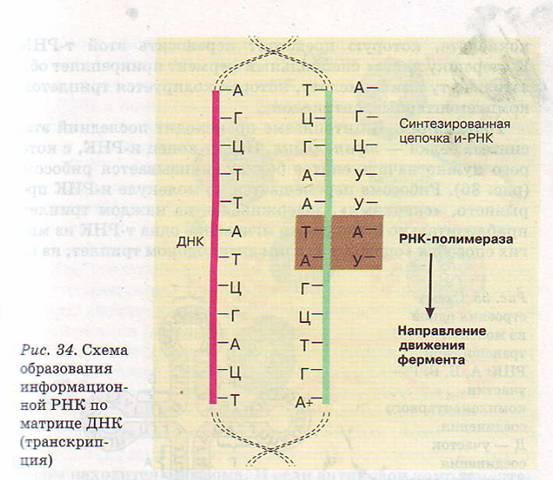
рис.1 Ну, например, для того чтобы синтезировать и-РНК, участок ДНК «разматывается», деспирализуется, а затем по принципу комплементарности на одной из цепочек ДНК с помощью ферментов синтезируются молекулы РНК (рис.1). "Группа Чукч..." правильно определила, что место "расплетения" является своеобразной рамкой для излучения ЭМ-полей, и центром сбора для тРНК в ядре клетке и инициатором движения созданных и нужных молекул.
рис.2 Рибосома так же является источником ЭМ-полей, и тот факт что она состоит из большой и малой субъединиц дает не сомневаться этому факту. Тем более, что тот факт что мРНК созданная в ядре должна под воздействием какой-то силы выйти из ядра и попасть к заданной рибосоме ( это явление тоже явно отличается от броуновского движения).
В обоих случаях "образ" инициализации движения сходен.
хоть биомолекулы и большие, излучать ЭМ-поля им нечем - мало свободных электронов, это вам не металлическая антенна на тв башне ; да и любое излучение может только биомолекулы накачать , что кодонов и АК не колышет, однако
А в отношении "переноса" аа-тРНК вы, в очередной раз, продемонстрировали свою бездарность, помноженную на безграмотность. В этом амплуа вы будете многие годы нудить и нудить тут одно и то же, "царствуя лежа на боку".
...Передвижение веществ внутри клетки осуществляется благодаря круговому движению цитоплазмы (циклозису) и направленной поперек этого движения диффузии...
Скорость этого движения не может восполнить скорость подачи нужных молекул в нужные места для сборки. Наличие этого движения (как второй производной) опосредовано и говорит о том, что есть внутренние инициаторы этого движения, о котором мы тут и говорим - это ЭМ-поля на АК и на РНК и места их прибытия.
Пользователю Хайдук... стоит напомнить наличие не большого количества электронов в клеточной структуре ПРОСТО говорит о слабых токах в ней, чайник вскипятить естественно явно не удастся.
ps Ну почему, когда говорят "розыгрыш призов", все слышат "призы" и никто не обращает внимания на "розыгрыш"?!
Оказывается и доцент П.Гаряев внёс свою лепту в доказательную часть «Группы Чукч..», ну например о целенаправленном круговом движении, правда он как обычно забыл сказать об источнике этого не стандартного движения, зря, так как он явно мог претендовать в соавторы на «Нобеля», об открытие дополнительного вида внутриклеточного транспорта для специфического обслуживания геномных регуляторов
ну вот видите, Петрович, как всё просто и НЕТ надобности в ваших сиомах, голографии и прочей фигне
При "водородных выдергиваниях" сиомия и её проблема никуда не деваются. Решение их остаются те же, что вы не поняли, точнее, не прочитали. Для 3-й виртуально вобулирующей буквы кодона срабатывает принцип "делегирования" перекодировочного смысла для этой буквы.
раньше предложил 3-4 таблицы с кодон-антикодон соответствиями, что решают все вопросы и для "вобулирующих сиомий" места НЕ остаётся. Period.
раньше предложил 3-4 таблицы с кодон-антикодон соответствиями, что решают все вопросы и для "вобулирующих сиомий" места НЕ остаётся.
Таблицы Кода стационарны, не учитывают факта реального чтения рибосомами текстов мРНК. Но даже в мертвой неподвижности Таблицы Кода видны дыры размером с Монблан. В эти дыры легко увидеть, что 3-й (3') нуклеотид не синонимов (сиом) одновременно ВИРТУАЛЬНО кодирует по две разных аминокислоты или аминокислоту и стоп позицию. А это для модели НЕ ПРИЕМЛЕМО. Это противоречие снимается контекстуальными ориентациями рибосомы на мРНК. Читай доказательства двойного кодирования в сатье Туранова и др.
Genetic Code Supports Targeted Insertion of
Two Amino Acids by One Codon
Anton A. Turanov, Alexey V. Lobanov, Dmitri E. Fomenko, Hilary G. Morrison,
Mitchell L. Sogin, Lawrence A. Klobutcher, Dolph L. Hatfield, Vadim N. Gladyshev*
*To whom correspondence should be addressed. E-mail:
Этот адрес электронной почты защищен от спам-ботов. У вас должен быть включен JavaScript для просмотра.
Published 9 January 2009, Science 323, 259 (2009)
DOI: 10.1126/science.1164748