Vladimirovich написал(а):
Отправлено в Лингвистико-Волновой геном (№16) Закрома.org
Оно, конечно, правильно - флуд в закрома. Но ведь это всего лишь продолжение темы ЛВГ, только в запредельном варианте.
Шумим-ругаемся. А главное-то остается: ген, конкретный ген, мы передали на расстояние нескольких см.. Пермиссивно передали - то есть включили в стволовых клетках реципиентах. Включили с помошью ген-информацией загруженного мШЭИ. И белок-маркер конкретный, свойственный только определенному типу клеток, был синтезирован. Вот где голову сморщить надо бы. И перестать браниться. Глупо. Итак, напомню это, главное:
Донорно-реципиентное дальнее волновое включение гена белка ххххх ткани мозга в культуре ххххххххх стволовых клеток хххххх
другой вариант
Донорно-реципиентное лазерно-радиоволновое включение гена ххххх в стволовых клетках ххххх
хххххх*., Гаряев П.П**., ххххххххх**., ххххххххххх**
*
*
*
** НП Новая биология 21 века
Материалы и методы. Биологическая часть.
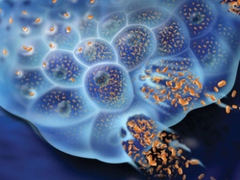
Мозг крысы линии Вистар, в соответствии с правилами гуманного обращения с лабораторными животными, после декапитации быстро извлекают, помещают в чашку Петри. хххх область коры головного мозга, как Донора волновой (электромагнитной) информации, сканируют лучом специального Гелий-Неонового лазера мощностью 2 мВт на расстоянии 10 см. В 20 см от облучаемого мозга и лазера (вне его луча) помещают флакон с монослойной культурой ххххххх стволовых клеток в среде _____. Воздействие на хххх осуществляют вторичным модулированном широкополосном электромагнитным излучением (мШЭИ) лазера [Прангишвили, Гаряев и др., 2000; Гаряев и др., 2007; Гаряев, 2009] в течение 1 часа. Затем культуру клеток перемещают в инкубатор при температуре хххххххх и продолжают культивирование в течение хххххххх со сменой культуральной среды хххххххх. В качестве контроля используют клетки, не подвергшиеся воздействию мШЭИ лазера. Через 3 недели опытные и контрольные образцы хххх фиксируют и проводят иммуногистохимическое исследование на наличие белка хххх - специфического маркера клеток ххххх [ ]. Для этого опытные и контрольные образцы окрашивают антителами против хххх. Эксперименты показали, что опытных образцах культур ххххх присутствуют клетки, иммунопозитивные к хххх (Рис.1). В культуре ххххх, не подвергшейся воздействию мШЭИ, клетки с хххххх не выявляются.
Физическая часть.
Для получения модулированного корой мозга широкополосного электромагнитного излучения (мШЭИ) применяли ранее разработанную нами биотехнологию работы с использованием специального Гелий-Неонового лазера [Гаряев, Тертышный, 1999]. Гелий-неоновый лазер мощностью 2 мВт и длиной волны 632.8 нм имеет две совмещенные, ортогональные линейно-поляризованные моды излучения, одночастотные в каждой из них. Лазерный луч зондирует препарат коры головного мозга крысы линии Вистар и при этом ортогональные поляризационные моды лазера модулируются в зависимости от способности зондируемых образцов поляризовать лазерный луч, вызывая специфическую перекачку энергии излучения между поляризационными модами. При этом модуляции оптических мод лазера связаны (коррелированы) с модуляциями электромагнитных векторов мШЭИ [Гаряев, 2009]. Существенным фактором такого зондирования является необходимость частичного отражения луча лазера обратно с резонатор лазера. Без такого обратного отражения луча, генерации мШЭИ не происходит и биологический эффект - запуск биосинтеза ххххх в ххххх - не наблюдается. Не менее важно, что такой встречный многопроходный режим луча позволяет зондируемому препарату выступать одновременно в роли оптического коррелятора [Мазур, Грачев, 1985] и донора волновой информации в режиме голографирования препарата на встречных лазерных пучках [Гаряев, 2009]. Оптические сигналы регистрируют и подают на электронную схему, которая управляет генерацией лазерного луча и мШЭИ. В таком режиме работы лазер генерирует, помимо красного света, еще и ШЭИ, модулированное препаратом, т. е. фактически само мШЭИ [Прангишвили, Гаряев и др., 2000]. Донором волновой информации в форме мШЭИ выступает препарат коры головного мозга крысы, в т.ч. клетки хххххх, а реципиентом информации - хххх. Важно, что культура клеток реципиентов мШЭИ располагается вне луча лазера и воспринимает генетическую информацию только от мШЭИ. Расстояние от зондируемого препарата до лазера составляет от 10 и более сантиметров.
Эксперимент
Предифференцировка и дифференцировка стволовых клеток ххххххххххххх – одно из ключевых направлений в клеточной биологии и медицине. ххххххххххххххххххххх кислотой [ ]. Способ заключается в индуцировании дифференциации хххххх стволовых клеток в ххххх клетки-предшественники путём культивирования ххххххх с плотностью ххххххххххх. хххххххххх в этом случае не исключена контаминация вирусами из фидерных клеток, которые используются на этапе получения клеточных агрегатов.
В нашем варианте предложен бесконтактный, волновой (электромагнитный) метод индукции клеточной дифференцировки, исключающий контаминацию патогенной флоры. Это способ бесконтактного квантового запуска начального этапа дифференциации культуры хххх в клетки ххххх с включением экспрессии гена белка ххххх. Применение данного метода индукции в 70-90% клеток ххххх иммуноморфологически выявляется маркер клеток хххх - белок ххххх.
Обсуждение.
В представленной работе мы не наблюдали финальной цитодифференцировки хххх в клетки хххх под влиянием несущего информацию мШЭИ. Возможно, это связано с точными условиями экспозиции хххх в поле мШЭИ, которые неизвестны. Время экспозиции хххх в мШЭИ - хххх. За это время клетки постепенно гибнут в первые минуты, но хромосомная ДНК, как главный источник волновой информации, сохраняется. Получен экспериментальный факт «переноса» волновой информации от донора, т.е. от гена-маркера хххх в реципиента, в хххх, и активация этого гена с последующим биосинтезом его продукта - белка хххх в реципиенте. Это не исключает возможность переноса в процессе эксперимента других генов от хххх к хххх. Для этого нужны соответствующие белковые маркеры. Возможно, мы имеем дело с начальным этапом дифференцировки хххх в хххх или с одним из многих этапов, запускаемых мШЭИ. Отметим, что более значимый эффект аналогичной работы получен нами ранее при волновом переносе морфогенетической информации от препаратов поджелудочной железы на крыс с вызванным искусственным, т.н. аллоксановым, диабетом. При этом наблюдается полная регенерация поджелудочных желез у больных животных с их последующим полным выздоровлением. Гистологический анализ показывает, что поджелудочные железы у опытных животных полностью регенерированы de novo. В контроле регенерация отсутствует [Гаряев и др., 2007; Гаряев, 2009]. В нашей ранней работе [Гаряев, 1994)], используя другой, разработанный нами, метод считывания волновой морфогенетической информации, также удалось передать управляющие электромагнитные сигналы на реципиента. В качестве донора волновой информации в этом случае были использованы головастики шпорцевой лягушки Xenopus laevis на определенной стадии развития, а реципиентом служила эктодерма ранней гаструлы (ЭРГ) той же Xenopus laevis. В результате тотипотентные клетки ЭРГ дифференцировались в полный набор нейральных и мезодермальных клеток и тканей. Нам не ясен точный механизм такого волнового «переноса и включения» гена ххххх, также как и в экспериментах с регенерацией поджелудочной железы и ЭРГ. Однако, теоретически обосновывая и предсказывая эффекты такого рода, мы высказали гипотезу о возможности пермиссивной телепортации генетической информации в таких экспериментах [Прангишвили, Гаряев и др., 2000; Gariaev et al., 2001]. Другие возможные квантовые механизмы обнаруженных процессов также широко обсуждаются. В том числе идея триады иных уровней генетического кодирования - а) лингвистического (реального текстового, не метафорического), б) голографического и в) квантово нелокального [Гаряев, 2009]. Полученные результаты подтверждают наши ранние исследования и не противоречат теоретическим моделям по волновой трансляции работающей морфогенетической информации [Гаряев и др., 2007; Гаряев и др., 2009; Прангишвили, Гаряев и др., 2000; Гаряев., Тертышный, Товмаш, 2007; Гаряев, Кокая, Леонова-Гаряева и др., 2007]. Лазерный волновой перенос одного из генов, участвующих в фотосинтезе у растений, также продемонстрирован, но с использованием существенно другой технологии [Казначеев, Трофимов, 2004]. Следует отметить, что реальные первые эксперименты по волновому переносу генетической информации начал Цзян Кань Джень [Jiang Kanzhen,1981]. Затем, наряду с нашими подтверждениями этого, появились и другие аналогичные работы [Бурлаков и др., 1999: Будаговский и др., 2001], технически по разному демонстрирующие факт существования и работы генетической информации в форме физических полей.
Цель и смысл наших и независимых экспериментов по электромагнитной трансляции работающей волновой генетической информации — еще одно доказательство реальности квантовых генетических функций хромосом, а также связанных с ними лингвистических векторов генома. Что означают полученные нами данные о «переносе» работающей волновой информации от донора, т.е. от гена белка-маркера ххххх, и факт активация этого гена с последующим биосинтезом его продукта - белка ххххх в реципиенте, т.е. в хххх. Эти результаты можно рассматривать как одну из вероятных моделей межклеточных волновых информационных процессов, основанных на знаковых когерентных фотонных излучениях хромосом [Popp, 2000]. В целом же функции генома можно гипотетически представить как работу квантового биокомпьютера, использующего три собственных, хромосомных, базовые атрибута - лингвистичность, голографичность и квантовую нелокальность [Gariaev et al., 2001; Гаряев, 2009]. Подчеркнем, что эти идеи лишь дополняют и развивают классическое, чисто вещественное, понимание функций генетического аппарата. Относительно лингвистичности (реальной текстовости, речеподобности ДНК - РНК) на уровне триплетного белкового кода: здесь с нами солидарен Л.П.Овчинников [
bio.fizteh.ru/student/files/biology/bioarticles/f_4ai2], по своему повторяющий нашу идею о контекстных (смысловых) ориентациях рибосом при считывании информационной РНК (мРНК). По мнению Л.П.Овчинникова это приводит к пониманию существования «второго генетического кода». Если вернуться к нашим данным по представленной работе, то мы имеем, по сути, известную проблему «черного ящика»: на входе - сигналы мШЭИ, вводимые в хххх. На выходе - активация в ххх гена хххх и биосинтез его белкового продукта. Как и в какой форме закодирована информация в мШЭИ о включении этого гена, мы можем только предполагать, давая несколько физико-математических версий запущенных квантовых и квази речевых событий в реципиентах [Гаряев, 2009]. Квази речевые (текстовые, смысловые) акты в белок синтезирующем аппарате хххх и в других биосистемах немыслимы без принятия идеи о геноме-биокомпьютере или ДНК компьютинге [Gariaev, 2001, Гаряев, 2009]. Такой биокомпьютинг может проходить на битовом цифровом, бинарно-счетном уровне [Shcherbak, 2003]. В.И.Щербак, анализируя количественные соотношения нуклонного состава ядер атомов кодируемых аминокислот и кодонов триплетного генетического кода, обосновал наличие системы арифметических операций в процессе биосинтеза белков, что также является проявлением некоторых сторон квази мышления генома. В.И.Щербак обнаружил в белковом коде систему генетического исчисления и использование ею функций ноля. Это чрезвычайно важное обстоятельство, поскольку ноль — сугубо мыслительное, запредельно абстрактное порождение, дающее начало координатному сознанию с его количественными мерами оценки внешнего мира. Эти оценки интерпретируются внутренним организменным генетическим сознанием-исчислением. Таким образом, цифры (наряду с буквами) становятся неотъемлемой частью генетического (белкового) кода. Поэтому арифметическое управление в лингвистической и/или текстовой генетике, полагает — В.И.Щербак, — реальность. Мы также видим квази разумность генома, но с другой, дополняющей, стороны - в его способности анализировать и понимать контекст мРНК и, как следствие, принимать точные решения в неопределенной ситуации выбора аминокислот и стоп позиций при считывании рибосомой не синонимических кодонов [Lagerkvist, 1978; Гаряев, 1997; Гаряев, 2009]. Существенным представляется, что геном, вероятно, может работать также и как квантовый биокомпьютер на кубитовом уровне. В этом смысле мШЭИ информация может передаваться квантово нелокальным путем [Gariaev, 2001; Гаряев, 2009], что совпадает с идеями Люка Монтанье и соавторов [ ]. В этом случае рибосомы могут работать как квантовые нанобиокомпьютеры [Тертышный, Гаряев, 2007, Гаряев, 2009]. В трактовке наших экспериментов нельзя также исключить особый феномен, связанный с так называемой фантомной памятью ДНК как возможным фактором передачи волновой генетической информации от донора к реципиенту как в данном случае, так и на межклеточном уровне in vivo. Это явление было обнаружено в 1984 году, опубликовано и объяснено [Gariaev et al., 1991; Гаряев, Тертышный, Товмаш, 2007; Gariaev, Pitkanen, 2010] и запатентовано [ патент на фантомы ]. Фактически аналогичное явление независимо обнаружено группой Р.Пекоры в Стэнфордском университете и названо «...mimicing the effect of dust» [Allison et al., 1990]. Трактовка этого феномена весьма трудна и требует привлечения не только принципов голографии, но и формализованных физико-математических понятий «темной материи» и «темных фотонов». Это дает одно из разумных объяснений как наших экспериментов, так и трансляцию структуры ДНК на воду группой Люка Монтанье [Gariaev, Pitkanen, 2010; Montagnier et al., 2010].
Литература
Гаряев П.П., Тертышный Г.Г. Приоритет на патент от 06.01.1999 г., № 99/01/Л, Федеральный институт промышленной собственности. Группа получающего ведомства РСТ. Старфилд, ЛТД.
Мазур А.И., Грачев В.Н. 1985. Электрохимические индикаторы. М., Радио и связь.
Прангишвили И.В., Гаряев П.П., Тертышный Г.Г., Максименко В.В., Мологин А.В., Леонова Е.А., Мулдашев Э.Р. 2000. Спектроскопия радиоволновых излучений локализованных фотонов: выход на квантово-нелокальные биоинформационные процессы. Датчики и Системы,
№ 9, Т. 18, с. 2-13.
Гаряев П.П., Прангишвили И.В., Тертышный Г.Г.,. Мологин А.В., Леонова Е.А., Мулдашев Э.Р. 2000. Генетические структуры как источник и приемник голографической информации.
Датчики и Системы, № 2, с. 2-8.
Гаряев П.П., Тертышный Г.Г., Товмаш А.В. 2007. Экспериментальные исследования in vitro по
голографическому отображению и переносу ДНК в комплексе с информацией, ее окружаю-
щей. Новые медицинские технологии, № 9, с. 42-53.
Гаряев П.П., 2009, Лингвистико-Волновой Геном. Теория и практика. Моногр. Киев. 220 с.
Гаряев П.П., Кокая А.А., Мухина И.В., Леонова-Гаряева Е.А., Кокая Н.Г. 2007. Влияние моду-
лированного биоструктурами электромагнитного излучения на течение аллоксанового са-
харного диабета у крыс. Бюллетень Экспериментальной Биологии и Медицины, № 2, с.155-
158.
Гаряев П.П., Кокая А.А., Леонова-Гаряева Е.А., Мулдашев Э.Р., Мухина И.В., Смелов М.В., Тертышный Г.Г., Товмаш А.В., Ягужинский Л.С. 2007. Теоретические модели волновой генетики и воспроизведение волнового иммунитета в эксперименте. Новые медицинские технологии, Новое медицинское оборудование, № 11, с. 26-70.
Gariaev P.P., Birshtein B.I., Iarochenko A.M., Marcer P.J., Tertishny G.G., Leonova K.A., Kaempf U. 2001. The DNA-wave biocomputer. «CASYS», International Journal of Computing Anticipatory Systems (ed. D.M.Dubois), Liege, Belgium, v. 10, pp. 290-310,
www.rialian.com/ rnboyd/dna-wave.doc
Jiang Kanzhen. 1981. The method to change organism’s heredity’s and the device to transmit
biological information. Soviet Union Patent № 1828665.
Popp F.A., in: Biophotonics and Coherent Systems. Proc., 2000, 118. 2-nd A.Gurwitsch Conf. and Add. Contrib. Eds by L.Beloussov, F.A.Popp, V.Voeikov, R.van Wijk. Moscow State University Press.
Shcherbak V.I. 2003. Arithmetic inside the universal genetic code. BioSystems, v. 70, pp.
187–209.
Lagerkvist U. 1978. «Two out of Three»: an alternative method for codon reading. Proc. Natl.
Acad. Sci., USA, v. 75, pp. 1759 1762.
Гаряев П.П.,1997, Волновой генетический код. Монография. М. 108с.
Тертышный Г.Г., Гаряев П.П. 2007. Волновые генетические нанотехнологии управления биосистемами. Теория и эксперименты. Новые медицинские технологии, № 7, с. 49-64.
Gariaev P.P., Chudin V.I., Komissarov G.G., Berezin A.A., Vasiliev A.A. 1991. Hologrphic Associative Memory of Biological Systems. Proceedings SPIE, The International Society for
Optical Engineering. Optical Memory and Neural Networks., USA, v. 1621, pp. 280-291.
Allison S.A., Sorlie S.S., Pecora R. 1990. Brownian Dynamics Simulations of Wormlike chains:
Dynamics Light Scattering from 2311 Base Pair DNA Fragments. Macromolecules, v. 23, pр.
1110-1118.
Гаряев П.П., Тертышный Г.Г., Товмаш А.В. 2007. Экспериментальные исследования in vitro по голографическому отображению и переносу ДНК в комплексе с информацией, ее окружающей. Новые медицинские технологии, № 9, с. 42-53.
Бурлаков А.Б., Бурлакова О.В., Голиченков В.А. 1999. Дистантные взаимодействия разново-
зрастных эмбрионов вьюна. ДАН, Т. 368, № 4, с. 562-564.
Будаговский А.В., Туровцева Н.И., Будаговский И.В. 2001. Когерентные электромагнитные
поля в дистанционном межклеточном взаимодействии. Биофизика. Т. 46, № 5, с. 894-900.
Технический аппендикс
Фиг.1.
Фиг.2.
ФИГ. 1. Блок- схема, реализующая донорно-реципиентное дальнее волновое включение гена белка BLBP радиальной глии в культуре мезенхимальных стволовых клеток костного мозга.
ФИГ. 2 . Блок-диаграмма донорно-реципиентного дальнего волнового включения гена белка BLBP радиальной глии в культуре мезенхимальных стволовых клеток костного мозга.
Технология дальнего волнового включения гена белка ххх в культуре хххх стволовых клеток хххх с помощью мШЭИ Гелий-Неонового лазера, в соответствии с фиг.1, характеризуется тем, что содержит фотодетектор 1, частотно-стабилизированный лазер 2, прозрачные стекла с донором 3, оптический фильтр 4, радиопрозрачный оптический затвор-шторка 5, кювету с акцептором 6, устройство поглощения обратного отражения лазерного света 7, блок обратной связи 8, радиоприемное устройство 9, компьютер 10, радиопередающее устройство 11, устройство излучения акустических волн 12.
Устройство работает следующим образом. См. Фиг. 2.
При возбуждении газового разряда в активном элементе лазера 1 между зеркалами 2 и 3 возникает генерация светового излучения на двух ортогонально поляризованных модах. Частота этого излучения определяется в основном расстояниям между зеркалами 2 и 3, но в некоторой степени и оптической длиной между зеркалами 3 и 6. Диэлектрическая трубка 4 позволяет изменять расстояние между зеркалами 2 и 3 Способом терморегулирования с использованием нагревателя 14. Этот Способ основан на том, что при изменении частоты излучения амплитуда ортогонально поляризованных мод изменяется в противофазе и это свойство используется для стабилизации частоты. Для этого излучение со стороны зеркала 2 разделяется по поляризациям с помощью поляризационного расщепителя 7 и подаётся на соответствующие площадки фотоприёмника 8. Сигналы фототоков вычитаются друг из друга в системе автоподстройки 11 и после соответствующего предварительного усиления подаются на усилитель мощности 12, который поддерживает такой ток в нагревательном элементе 14, чтобы интенсивность ортогонально поляризованных мод в среднем была одинаковой. При этом, если температура окружающей среды растёт, то уровень подачи энергии на нагреватель 14 уменьшается, и наоборот, если температура окружающей среды падает, то уровень подачи энергии на нагреватель 14 растёт. Тем самым осуществляется стабилизация средней частоты излучения. Использование дополнительной отражающей поверхности, расположенной в оптическом фильтре 6, и отраженному от нее свету, который прошел донор, и возвращен обратно в лазерный резонатор, приводит к созданию в области донора, заключенного в кювету или расположенного между прозрачными стеклами 5, двух оптических решеток из стоячих волн ортогонально поляризованных мод. Причём фазовый сдвиг отраженного от зеркала излучения зависит от поляризационных и других физико-информационных свойств донора (например, поджелудочной железы), расположенного между прозрачными стеклами 5. В результате суммарное пропускания зеркал 3 и 6 изменяется, и излучение лазера флуктуирует по частоте и амплитуде. Уровень флуктуации зависит от угла нормали частичного отражателя 6 к оптической оси и динамических параметров донора, находящегося в кювете 5. Эти колебания приводят к изменениям в структуре стоячих волн в обеих модах с ортогональными поляризациями. Флуктуации амплитуд ортогонально поляризованных мод приводят к флуктуации тока, подаваемого на фотодетектор и далее через систему обратной связи на нагреватель 14 и, соответственно, к флуктуациям уровня мощности, потребляемого усилителем мощности 12 от импульсного источника 13. Эти флуктуации потребляемой мощности приводят к флуктуациям частоты генератора импульсного источника. Они и являются широко спектральным радио волновым (мШЭИ) 15 и акустическим 16 источником программирующих реципиент излучений. Эти излучения содержат волновую информацию, считанную с донора, размещенного на предметном столике или между прозрачными стеклами 5.
При соответствующей настройке и калибровке оптических элементов уровни и частоты оптических флуктуации лазерных мод коррелированы с флуктуациями уровней и частот в излучаемом акустическом и радиоволновом (мШЭИ) диапазоне волн. Эти волны, излучаемые радио волновым источником 15 и акустическим излучателем 16, используются для воздействия донорной информацией на клетки реципиента хххх. В данной работе использовали только радио волновой источник информации (мШЭИ), считанной с донора, т. е. с хххххх. Это достигается посредством облучения мШЭИ реципиента 10 при радиопрозрачном затворе 9 в течении от хххх и более хххх в зависимости от состояний донора и реципиента и поставленной задачи относительно волнового мШЭИ воздействия.
Управляющее воздействие по каналу мШЭИ происходит при экспонировании реципиента на расстоянии от сантиметров до многих метров от лазера и вне его луча.