TOPIC: Лингвистико-Волновой геном. Следующий шаг. №3
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 13:03 #61
PauLita написал(а):
Продолжением или нет, но он до сих пор неопубликован. Что уж говорить об рецензировании или подтверждении!
Так это то же что и в постах 29 и 42. По большому счету ничего публиковать не нужно. Более того в статье в постах 29 и 42 гораздо более крутой эксперимент чем по вибропортациии. Эксперимент по вибропортации всего лишь подмножество и жалкое подобие того что было в январе этого года. А статья в постах 29 и 42 прорецензирована
PauLita написал(а):
А в указанном Вами материале лишь говориться, что от чистой воды регистрируется такой же ЭМ сигнал, как и от грязной. От этого до вибропортации еще ой как далеко
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 13:16 #65
Статья Electromagnetic signals are produced by aqueous nanostructures derived from bacterial DNA sequences прорецензирована, но в ней отсутвует вибропортация ДНК. Статья arXiv:1012.5166v1 [q-bio.OT] 23 Dec 2010 DNA waves and water никем не рецензированна. По приведенному в ней беглому описанию эксперимента повторить его невозможно. Несмотря на обещание Люка, статья о вибропортации ДНК до сих пор не опубликована ни в одном рецензируемом журнале. Прошло уже полгода!
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 13:19 #66
PauLita написал(а):
но в ней отсутВует вибропортация ДНК.
Не отсутВует а - присутВует. Только не искусственно созданная а природная . Иначе бы микоплазма в стерильной воде не появлялась. И ПЦР там та же что и в препринте. Да Вы почитайте тот фрагмент что я здесь запостил из препринта - там написано что отталкивались от той прорецензированной публикации.
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 13:30 #67
limarodessa написал(а):
Не отсутВует а - присутВует. Только не искусственно созданная а природная . Иначе бы микоплазма в стерильной воде не появлялась. И ПЦР там та же что и в препринте. Да Вы почитайте тот фрагмент что я здесь запостил из препринта - там написано что отталкивались от той прорецензированной публикации.
И так, в первом эксперименте, PCR применяют для проверки исходного материала. Во втором - только в конце, для доказательства того, что ДНК была вибропортирована. Ну а Вы делаете совершенно необоснованный вывод, что и во втором эксперименте применялись те же условия, что и в первом.
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 13:31 #68
PauLita написал(а):
И так, в первом эксперименте, PCR применяют только в начале для проверки исходного материала. Во втором - только в конце, для доказательства того, что ДНК была вибропортирована. Ну а Вы делаете совершенно необоснованный вывод, что и во втором эксперименте применялись те же условия, что и в первом.
Да потому что в первом была микоплазма, ее может и иначе зарегистрировать можно. А во втором там ДНК поэтому ПЦР. Но сути это аж никак не меняет. ДНК микоплазмы тоже была вибропортирована но только природным механизмом а не катушкой индуктивности. Поэтому мой вывод совершенно обоснован
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 13:41 #69
limarodessa написал(а):
Поэтому мой вывод совершенно обоснован
Как только Люк опубликует свой эксперимент в рецензируемом журнале, а потом кто нибудь еще его повторит, вот тогда и только тогда можно будет говорить об обоснованности. Причем не в журнале Interdiscip Sci Comput Life Sci типа сам пишу, сам рецензирую
Editor-in-Chief:
Dongqing Wei, Shanghai Jiaotong University, China
Executive Editor
Limin Angela Liu, Fred Hutchinson Cancer Research Center and University of Washington, USA
Associate Editors
Joel S. Bader, Johns Hopkins University, USA
Liqun Chen, Shanghai University, Shanghai, China
Zizheng Gong, China Academy of Space Technology, Beijing, China
Hong Guo, University of Tennessee, USA
Zengjian Hu, Howard University, USA
Igor Jurisica, University of Toronto, Canada
Shaokai Luo, Zhejiang Sci-Tech University, Hangzhou, China
Jean-Philip Piquemal, Universit Pierre et Marie Curie, France
Qiushi Ren, Shanghai Jiaotong University, China
Russell Schwartz, Carnegie Mellon University, USA
Qingming Zhang, Beijing Institute of Technology, China
Editorial Board
Chairman
Luc Montagnier, World Foundation AIDS Research and Prevention, France
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 13:57 #70
PauLita написал(а):
Cell and Molecular Biology, 2003 написал(а):
There is no evidence to indicate that genetic code is ambiguous in vivo. Ambiguity means that a single codon may code for more than one amino acid. The only exception to the non ambiguity is the AUG codon in prokaryotes – it codes for formyl methionine at the initiation site while at other positions it specifies methionine. It is not clear as to how this distinction is always met precisely but it is believed that the secondary structure of the mRNA at the initiation site may havgot something to do with the coding for formyl methionine. In in vitro conditions however genetic codes sometime appear to be ambiguos. For example poly-U codon is known to incorporate some amounts of leucine in addition to phenylalanine. Treatment of ribosomes with streptomycin is known to introduce ambiguity to codons. For example in addition to coding for phenylalanine UUU is also known to code isoleucine, serine and leucina.
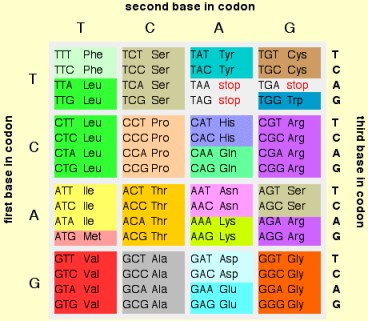
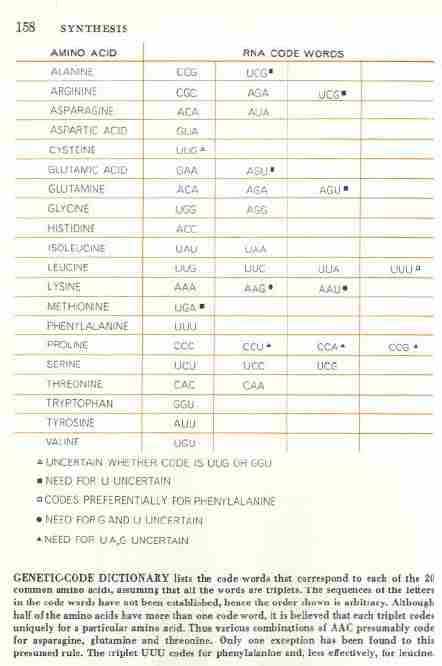
Любезный, этой цитатой вы сами себя и высекли. Она как раз и подтверждает идею неоднозначного кодироания половины кодонов. А вторая цитата пытается это замазать. Еще раз, Ниренберг и Крик видели это противоречие уже в UUU кодоне и признали это. Я месяцами искал оригинал этого перевода. Нет его нигде. Почему? Да потому, что позор - нобеля дали за ошибочную модель кода Ниренбергу в 1968г. Найдете оригинал - дайте ссылку. Единственный, кто после меня признал фундаментальность контекстных ориентаций в рибосомальном синтезе - это акад. РАН Л.Овчинников. И назвал это ВТОРЫМ ГЕНЕТИЧЕСКИМ КОДОМ. Крик перед смертью чечтно признал, что не видит ОЧЕВИДНОГО смысла в модели кода. А неочевидный в омонимических двусмысленностях кодонов-омонимов.
Попутно отвечу и PP. Сиквенсы белков предлагает делать. Для чего? Как сиквенсами (полученными последовательностями аминокислот в белках) доказать неправомочность идеи мРНК контекстных ориентаций при синтезе белков? Ответить не может и не ответит никогда.
Поясню на примере, если прямая логика математику недоступна. Допустим, я задаю PP вопрос: вы слова 'ключ' или 'лук' (омонимы) как понимаете? Естественно, он ответит, что значение, смысл этих слов, как частей фразы или предложения, зависит от общего смысла фразы или предложения. От контекста. И будет абсолютно прав. А я ему - а докажите-ка это... Это совершенно аналогично тому, что предлагает мне PP - доказать, что рибосома понимает значение кодонов-омонимов только в контексте мРНК. Как вы, любезный PP, экспериметально докажете контекстуальную роль для точного понимании слов 'ключ' или 'лук'? Ась... не слышу ответа... Доказывать тут ничего не надо в виду очевидности ситуации. С рибосомами не так все очевидно, возразит мне PP. И будет тоже прав. Как доказать для рибосом, я дал в моей 3-й монографии. Можно сделать также модельные синтезы белков с cell free system и известными последовательнстями РНК. Вы практически всегда получите нормальные белки, последовательность аминокислот которых будут отражать РНК контекстные влияния. А если уж совсем глубоко копать, то вы должны будете доказать, что белок синтезирующий аппарат обладает способностью к элементарному мышлению, т.е. это нанобиокомпьютер, если угодно и удобно так считать.
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 14:10 #71
ППГ написал(а):
Крик перед смертью чечтно признал, что не видит ОЧЕВИДНОГО смысла в модели кода.
Очередная ложь! Вот оригинал:
Francis Crick. What Mad Pursuit. A Personal View of Scientific Discovery написал(а):
An important point to notice is that although the genetic code has certain regularities – in several cases it is the first two bases that encode one amino acid, the nature of the third being irrelevant – its structure otherwise makes no obvious sence. It could well be that it is mainly the result of historical accidents in the distant past.
...
This is a nice example of complexity of nature produced by natural selection. It shows how easily one can be misled if one takes too straightforward a view of biological problem. Of course, we were fortunate to have hit on the correct standard set at our first attempt. It was lucky guess and needed to be confirmed by many additional experiments. While it took some years for biochemists to do this, that our list was correct was never seriously in doubt. Although there was occasional confilicting evidence, our list has stood the test of time.
...
After I returned to England in the fall of 1956 I wrote a paper for it analyzing Gamow‘s ideas, generalizing them, and suggesting what turned out to be an important idea, the adaptor hypothesis.
The paper was called „On Degenerate Templates and the Adaptor Hypothesis.“ The main idea was that it was very difficult to consider how DNA or RNA, in any conceivable form, could provide a direct template for the side-chains of twenty standard amino acids. What any structure was likely to have was a specific pattern of atomic groups that could form hydrogenic bounds. I therefore proposes a theory in which there were twenty adaptors (one for for each amino acid), together with twenty special enzymes. Each enzyme would join one particular amino acis to its own special adaptor. This combination would then diffuse to the RNA template. An adaptor molecule could fit in only those places on the nucleic acid template where it could form the necessary hydrogen bonds to hold it in place. Sitting there, it would have carried its amino acid to just the right place it was needed.
There were several implications of this idea. The one I want to stress here was that it meant that genetic code could have almost any structure, since its details would depend on which amino acid went with which adaptor. This had probaly been decided very early in evolution and possibly by chance. Because of this pessimistic conclusion the paper led off with a quotation from an obscure Persian writer of the eleventh century: “Is there anyone so utterly lost as he that seeks a way where is no way?“ and ended with the remark, „In the comparative isoltion of Cambridge, I must confess there are times when I have no stomach for the coding problem.“
Специально для тебя, Петя, еще и еще раз повторяю: Не видит очевидного смысла не в модели кода, а в структуре [таблицы] кода
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 14:17 #72
PauLita написал(а):
Как только Люк опубликует свой эксперимент в рецензируемом журнале, а потом кто нибудь еще его повторит, вот тогда и только тогда можно будет говорить об обоснованности.
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 14:26 #74
PauLita написал(а):
Статья Electromagnetic signals are produced by aqueous nanostructures derived from bacterial DNA sequences прорецензирована, но в ней отсутвует вибропортация ДНК. Статья arXiv:1012.5166v1 [q-bio.OT] 23 Dec 2010 DNA waves and water никем не рецензированна. По приведенному в ней беглому описанию эксперимента повторить его невозможно. Несмотря на обещание Люка, статья о вибропортации ДНК до сих пор не опубликована ни в одном рецензируемом журнале. Прошло уже полгода!
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 14:40 #77
PauLita написал(а):
Специально для тебя, Петя, еще и еще раз повторяю: Не видит очевидного смысла не в модели кода, а в структуре [таблицы] кода
Вот правильно. Еще раз сам себя высек, любезный. Структура (таблица) кода просто кричит, что 32 кодона НЕОДНОЗНАЧНЫ. а ПОСЕМУ И САМА МОДЕЛЬ ЛОЖНА. Л.Овчинников, классик рибосомологии (вместе со А.Спириным), однозначно говорит о ВТОРОМ генетическом коде, основанном на контекстуальных эффектах при синтезе белков. Ошибка модели уже ДВАЖДЫ дорого обошлась человечеству в силу трансгенной безграмотности инженеров по ГМ продуктам питания и синей чумой в Мексиканском заливе с ее пожирющими людей и все живое синтиями болвана крэйга вентера.
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 14:51 #80
Болван — деревянная форма, подставка с круглым верхом для расправления шляп, чепцов, париков (Ушаков).
Призываю воздерживаться от оффтопа! Вместо того искать конкретные и действенные аргументы против позиции оппонента, но нейтральные по отношению к личности его. (Хайдук)
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 15:11 #81
Vladimirovich написал(а):
Фрактал есть структура, метрическая размерность которой больше, чем размерность Лебега.
Что вкладывается в понятие фрактальность времени?
ФШ любой системы, состоит из одинакового количества временных интервалов, максимально из 5-7. Максимальное время всех процессов упорядочения энергии, задано экстенсивным параметром системы, её массой. Инвариантность структуры времени, наряду с явлением перераспределения времени от ведущего, наиболее устойчивого процесса к процессам менее устойчивым, указывает на фрактальность структуры времени. Фрактальность структуры времени определяет, вероятно, фрактальность распределения энергии в виде 5-7 потоков при переходе неравновесной системы в состояние, в котором она оказывает минимальное воздействие на окружающую среду. Благодаря инвариантности структуры времени и фрактальности распределения энергии в виде тонкой структуры гистограмм МФ, возможно такое изменение масштаба величины различных по составу и массе систем, которое обеспечивает совпадение формы гистограмм МФ различных процессов в различных системах, что отмечено в работе С.Э. Шноля и его коллег. www.chronos.msu.ru/RREPORTS/vanyarkho_metod/vanyarkho_metod.htm
Я понимаю фрактальность времени как некое древо (классический фрактал ветвления), где всё древо - вселенское время. Оно имеет ствол и ветвящиеся ветви, каждая из которых является самоподобной частью этого фрактала, подобная целой структуре (самоподобие или разномасштабная самотождественность). Толстые ветви - времена галалактик, тоньше - времена звезд, еще тоньше - времена планет, самые тонкие - времена жизни биосистем, их метаболических процессов. Самые тонкие - времена жизни элементарных частиц. Каждая веточка живет в масштабах общего вселенского времени, и вместе с тем, ей свойственна ее собственная длительнсть бытия. Фракктал, нелинейный, как нелинейно время по Эйнштейну. Интуитивно чувствую, что в хромосомном аппарате, как операторе биовремни, эти нелинейности и кривизны времени играют существенную роль для биосистем. Какую? Думаю. Интерсно увязать это с излучениями фотонов молекулами ДНК, релятивистские скорости их, замедляемые структурой биосистем, возможно связаны с течением времни в пространстве биосистем. Есть и еще соображения, в частности, те, о которых говорил пару лет назад и сейчас. Это о возможных холодных временнЫх фракталах, спасающих бактерий термофилов от высоких, несовместимых с жизнью, температур. В нас - это обеспечение когерентности биохимических процессов и возможность информационного спутывания. Сюда же относятся феномены прогерии (стремительного сстарения) и наоборот, торможения старения при летаргии, выживание организмов при замерзании и т.д. Но все, понятное дело, пока еще сыровато. Думать дальше надо.
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 15:20 #83
ППГ написал(а):
ФШ любой системы, состоит из одинакового количества временных интервалов, максимально из 5-7. Максимальное время всех процессов упорядочения энергии, задано экстенсивным параметром системы, её массой. Инвариантность структуры времени, наряду с явлением перераспределения времени от ведущего, наиболее устойчивого процесса к процессам менее устойчивым, указывает на фрактальность структуры времени. Фрактальность структуры времени определяет, вероятно, фрактальность распределения энергии в виде 5-7 потоков при переходе неравновесной системы в состояние, в котором она оказывает минимальное воздействие на окружающую среду. Благодаря инвариантности структуры времени и фрактальности распределения энергии в виде тонкой структуры гистограмм МФ, возможно такое изменение масштаба величины различных по составу и массе систем, которое обеспечивает совпадение формы гистограмм МФ различных процессов в различных системах, что отмечено в работе С.Э. Шноля и его коллег.
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 15:22 #85
PauLita написал(а):
Не видит [Крик] очевидного смысла не в модели кода, а в структуре [таблицы] кода
Придираться к словам не стоит. Лучше показать на конкретных кодонах/триплетах, что критика Петровичем модели/структуры кода Криком скорее безопасна для Фрэнсиса. К сожалению, Петрович НЕ конкретен и не указывает на специфический кодон, вызывающий синтез двух или более аминокислот в различных условиях. Если это действительно происходит, то очевидно надо будет посмотреть в чем состоит уникальность условий для каждой аминокислоты и сохраняется ли эта уникальность каждый раз, когда наш кодон (то бишь не наш, а Петровича
Лингвистико-Волновой геном. Следующий шаг. №3
01 Июль 2011 15:24 #86
ППГ написал(а):
Я понимаю фрактальность времени как некое древо (классический фрактал ветвления), где всё древо - вселенское время. Оно имеет ствол и ветвящиеся ветви, каждая из которых является самоподобной частью этого фрактала, подобная целой структуре (самоподобие или разномасштабная самотождественность). Толстые ветви - времена галалактик, тоньше - времена звезд, еще тоньше - времена планет, самые тонкие - времена жизни биосистем, их метаболических процессов. Самые тонкие - времена жизни элементарных частиц. Каждая веточка живет в масштабах общего вселенского времени, и вместе с тем, ей свойственна ее собственная длительнсть бытия. Фракктал, нелинейный, как нелинейно время по Эйнштейну. Интуитивно чувствую, что в хромосомном аппарате, как операторе биовремни, эти нелинейности и кривизны времени играют существенную роль для биосистем. Какую? Думаю. Интерсно увязать это с излучениями фотонов молекулами ДНК, релятивистские скорости их, замедляемые структурой биосистем, возможно связаны с течением времни в пространстве биосистем. Есть и еще соображения, в частности, те, о которых говорил пару лет назад и сейчас. Это о возможных холодных временнЫх фракталах, спасающих бактерий термофилов от высоких, несовместимых с жизнью, температур. В нас - это обеспечение когерентности биохимических процессов и возможность информационного спутывания. Сюда же относятся феномены прогерии (стремительного сстарения) и наоборот, торможения старения при летаргии, выживание организмов при замерзании и т.д. Но все, понятное дело, пока еще сыровато. Думать дальше надо.
Давайте... Вспомним гипотезу (теорию) градиентов морфогенов Вольперта и энтелехий Аристотеля-Дриша составляющих последовательности Ву-Кэбота. Она хорошо объясняет «синдром Крейцфельда-Якоба», «синдром Гертсмана-Штраусслера-Шейнкера» и «синдром Альперса“, а также спектр резонансов Ферми-Паста-Улама, промодулированных предельным случаем омонимии, известным в лингвистике и теории информации (теорема Шеннона). При этом эффект, имитирующий пыль следует фоново-ассоциативному принципу, который включает шенновский эффект как частный случай парадокса или канала Эйнштейна-Подольского-Розена (ЭПР). Распределение Ципфа-Мандельброта для частот на основе явления Ганса-Отто-Мейера сопровождается возникновением изоморфных временных автокорреляционных функций на так называемых Фрелиховских модах передающих динамику углов Эйлера вращающих плоскость поляризации проходящих через них оптических излучений. Фурье-образ радиоспектров мШЭИ дополняет известную модель Фрелиха возникающую при Бозе-конденсации фононов электромагнитных волн белков. При этом Давыдовский гамильтониан трансформирован в нормальные координаты, а Фрелиховский гамильтониан канонически трансформирован в эквивалентную форму в рамках аппроксимации Хартри-Фока. Структура преобразования связана с прямым и обратным Фурье-преобразованием, которое состоит, во-первых, в формировании и регистрации от каждой точки квазиколец Ньютона и, во-вторых, в их обратном Фурье-преобразовании многочленами Жегалкина вычисленными индуцированным отображением сдвига тора на тайлах разбиений Рози. В случае частичного стирания или вибрационного размытия квазиколец Ньютона, которые соответствуют некоторой точке, оставшаяся часть колец оказывается необходимой и достаточной для правильного формирования алгоритмом Штрассена для булевых матриц соответствующей точки и является генератором ФПУ-солитонных акустико-электромагнитных полей трансформируемых в совокупность объемных поляризационных конусов стоячей световой волны интенсивности.
Ув. Паулита, такое флудерьё неконструктивно, лучше сосредоточиться на критику своими словами возможной мифологии, если таковую застукали в изложении оппонента